Ewolucyjne kompromisy | Szlaki wzrokowe | Teorie percepcji wzrokowej | Złudzenia wzrokowe | Agnozje i zaburzenia wzrokowe | Wyobraźnia wzrokowa | Mechanizm widzenia
Zadaniem percepcji jest dostarczenie informacji o stanie ciała i jego otoczenia przetworzonej w taki sposób, by można było zareagować w przydatny sposób. Potrzebne są do tego odpowiednie receptory i podstawowe mechanizmy analizujące przesyłane przez nie informacje. W tej części zajmiemy się mechanizmami widzenia i kontroli ruchów.
B17.1 Wzrok i ewolucyjne kompromisy
Zmysły nie dostarczają nam informacji o obiektywnej rzeczywistości. Istnieje mit o "postrzeganiu świata jakim naprawdę jest". Ma to tylko częściowy sens z kilku przyczyn.
Jak omawialiśmy to na pierwszym wykładzie informacje zmysłowe pomagają nam podejmować decyzje, nie ma więc potrzeby postrzegania wszystkiego. Co jest dla nas istotne? Postrzeganie i kategoryzacja obiektów: kształt i kolor pomaga rozpoznać rzeczy jadalne i niejadalne, ruch pozwala dostrzec niebezpieczeństwo i potencjalną ofiarę.
Nie mamy możliwości eksploracji świata w obiektywny sposób, widziane światło o długości fali 380-750 nm to bardzo mały ułamek całego spektrum fal elektromagnetycznych, słyszane wibracje 20Hz-20kHz również mały fragment możliwych wibracji. Odczuwamy więc niezwykle mało, wiele zwierząt ma zmysły odbierające informację, której my nie dostrzegamy.
Na poziomie analizy sygnałów z pozostałych zmysłów również nie jest lepiej. Ewolucja ukrywa przed nami prawdę o fizycznych aspektach docierających do nas bodźców, bo nie są one nam przydatne. Widać to dobrze na przykładzie omówionych niżej iluzji wzrokowych i iluzji związanych z działaniem innych zmysłów. Odczuwamy tyle, ile nam się może przydać. Zbyt wiele informacji spowolni nasze decyzje i niepotrzebnie obciąży nasze mózgi. Modele matematyczne ewolucji funkcji pokazują, że organizmy nastawione na postrzeganie prawdziwych, fizycznych cech za pomocą zmysłów nie mają szans w konkurencji z organizmami, które postrzegają tylko informacje przydatne do działania (Hoffman, 2019). Ewolucja minimalizuje ilość docierających do nas informacji optymalizując ich przetwarzanie tak, by umożliwić nam podejmowanie decyzji sprzyjających przetrwaniu. Tak przynajmniej było do czasu, gdy człowiek w ciągu ostatnich paru tysięcy lat przekształcił swoje środowisko, nie dając ewolucji czasu by dostosować nasze organizmy do nowej sytuacji. W efekcie mamy liczne choroby cywilizacyjne.
Ponieważ nasze postrzeganie realizuje pewien proces dostarczania użytecznych informacji włączają się tu mechanizmy uwagi, aktywnego poszukiwania ważnych bodźców. Wpływ nastawienia, oczekiwań na to, co postrzegamy jest silny i bardzo przydatny przy wykonywaniu określonych czynności. W efekcie nie dostrzegamy wielu innych, nie potrafimy dostrzegać piękna krajobrazu, przyjemności muzyki czy jedzenia.
Jeśli uda się nam postrzegać świat bez żadnych uprzedzeń to struktura naszego mózgu i receptorów zmysłowych wcale nie pozwoli nam go widzieć "jaki jest naprawdę", ale będziemy mogli postrzegać świat mniej wybiórczo, a bardziej całościowo.
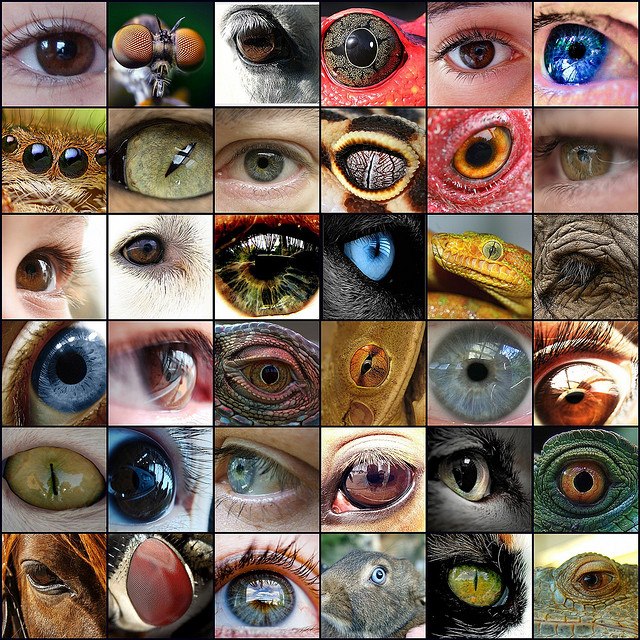
Wzrok pozwala dostrzec niebezpieczeństwo lub okazję do polowania z daleka, rozpoznać wroga lub ofiarę, dostrzec dojrzały owoc wysoko na drzewie, jest więc szczególnie cennym zmysłem.
Stare, mało precyzyjne mechanizmy związane są z ośrodkami podkorowymi, szybkimi reakcjami emocjonalnymi, mobilizacją organizmu i bezwarunkowymi odruchami refleksyjnymi.
Kolor czy ruch są perceptami, a nie tylko fizycznymi własnościami fal elektromagnetycznych czy akustycznych docierających do receptorów oka.
To oznacza, że jest to wysoce przetworzona informacja, przedstawiona w taki sposób by ułatwić proces podejmowania decyzji, reakcji organizmu na to co się dzieje w otoczeniu i co się dzieje z własnym ciałem. Przyrządy fizyczne analizujące widmo światła oprócz receptorów reagujących na światło też zawierają obwody analizujące stan tych receptorów i przesyłające wyniki do dalszego wykorzystania. Dlatego już na poziomie samej konstrukcji oczu odbywa się skomplikowane przetwarzanie sygnału przez różnego typu neurony.
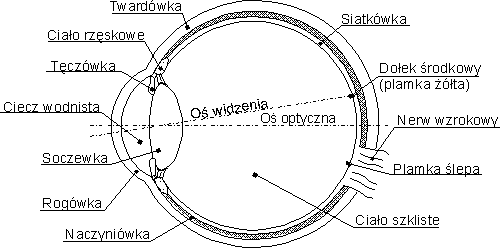
|
Wzrok u zwierząt różnego gatunku realizowany jest na wiele sposobów: ślimak ma komórki światłoczułe bez soczewek,
owady złożone oko i 10-30.000 heksagonalnych fasetek,
ssaki mają
oko z siatkówką i soczewką,
głowonogi mają oczy podobne do ssaków.
Oko jest wysuniętą na zewnątrz częścią mózgu, "zwierciadłem duszy".
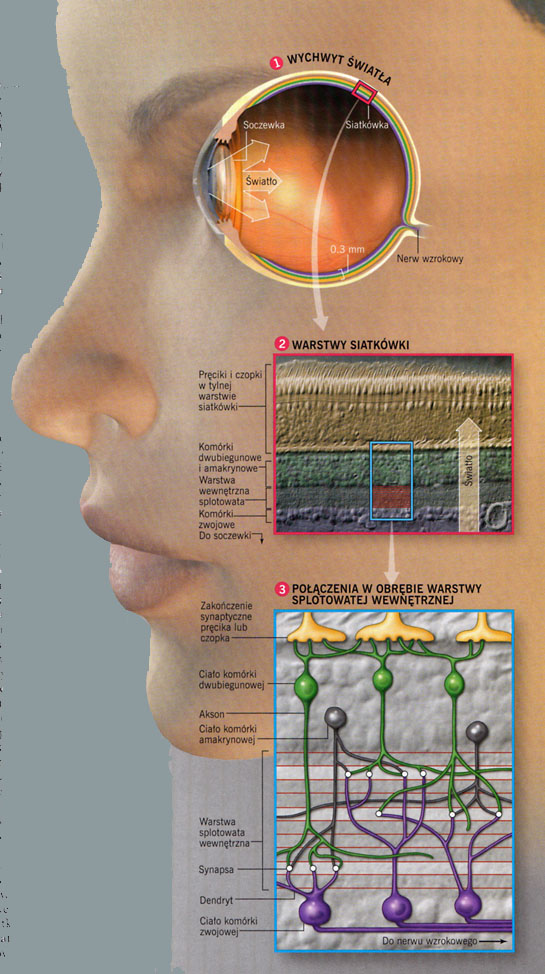
|
Oczy wykorzystują światło, powinny więc wyodrębniać ważne informacje; potrzebne są różne kompromisy:
- widzieć dobrze w ciemności jak i przy jaskrawym świetle - pomaga tęczówka regulując wielkość źrenicy (to działa jak przysłona w aparacie), ale dla silniejszych różnic nie da się tego zrobić jednym systemem i potrzebne są różne typy receptorów, które muszą się przełączać;
- widzieć ostro i szeroko - to wymaga szybkich ruchów gałki ocznej;
- reagować szybko i analizować dokładnie - to wymaga różnych metod transmisji informacji z siatkówki;
- widzieć we wszystkich kierunkach i spostrzegać głębię - do tego potrzebujemy więcej niż pary oczu, lub niezależnie poruszanych oczu jak kameleony;
- widzieć ten sam kolor niezależnie od barwy oświetlającego światła (południe, zachód Słońca) - to zadanie dla kory wzrokowej;
- widzieć blisko i daleko, umożliwić precyzyjne działanie i planowanie;
- widzieć stabilnie pomimo różnych zakłóceń, łzawienia, padającego deszczu, mgły czy zamieci.
Książka Webvision bardzo szczegółowo opisuje budowę oka, fotoreceptorów, siatkówki i trochę mniej dokładnie przetwarzanie informacji w korze wzrokowej.
Są w niej dane liczbowe na temat
siatkówki i jej połączeń. Struktura siatkówki stabilizuje się dopiero około 4 roku życia.
Nie będziemy tego dokładnie omawiać.
Siatkówka ma około pół milimetra grubości, niezbyt doskonałą złożoną budowę:
- światło musi najpierw przeniknąć przez trzy warstwy komórek neuronalnych,
- nerw wzrokowy, czyli wiązka aksonów przesyłających informację do kory mózgu, musi wyjść na zewnątrz przez plamkę ślepą,
- pręciki i czopki są w jej tylnej warstwie.
Fotoreceptory przesyłają impulsy do komórek dwubiegunowych, których jest aż 9 rodzajów, a te pobudzają komórki horyzontalne i komórki zwojowe (20 typów), oraz komórki amakrynowe (22 typy, lokalnie hamujące), których aksony tworzą nerw wzrokowy, wyprowadzający informację z siatkówki przez plamkę ślepą. Najważniejszym neurotransmiterem w tych warstwach jest kwas glutaminowy (glutaminian).
Tak skomplikowana budowa sprzyja powstawaniu licznych chorób siatkówki. W skali globalnej ponad 1/4 ludzkości jest
niewidoma lub niedowidzi. Oko to cudowny narząd, ale jak widać daleki od idealnej konstrukcji. Jest bardzo wiele chorób oczu.
Kompromis pomiędzy ostrym widzeniem pod szerokim kątem a szybkim i oszczędnym przetwarzaniem informacji wzrokowej został rozwiązany przez ewolucję na wiele sposobów, stąd mamy tyle różnych rodzajów oczu i mechanizmów ich wykorzystywania. Komórki światłoczułe mogą być rozrzucone po całym ciele, pole widzenia może być szerokie ale widzenie niewyraźne (pająki mają parę oczu i 6 pomocniczych, nosorożce słabo widzą), każde oko może się niezależnie poruszać (kameleony, ustonogi), sowa może obracać głowę o 270 stopni, a ssaki mają skomplikowany mechanizm określający kierunek spojrzenia. Bez przerwy wykonujemy drobne ruchy oczu by widzieć wyraźnie, a nasze widzenie peryferyjne i sygnały z innych zmysłów pozwalają nam zauważyć, gdzie należy skierować wzrok.
|
Co właściwie widzimy? By oszczędzać energię mózgi uczą się ignorowania tego, co jest nieistotne, co się nie zmienia. Gady nie widziały nieruchomych ofiar, stąd powszechny odruch zamierania u owadów, gadów i innych zwierząt. Już Arystoteles zauważył, że patrząc długo na płynącą wodę, a potem na skałę, mamy wrażenie, jakby to skała zaczęła płynąć. To znana "iluzja wodospadu". Mózg szybko przyzwyczaja się do tła, więc jeśli ciągły ruch staje się tłem zaczynamy dostrzegać tylko zmiany tego ruchu, a nie sam ruch. Powstaje proces kompensacji wrażenia ruchu, dlatego przy obserwacji nieruchomych obiektów przez jakiś czas mechanizm kompensacji stwarza wrażenie ruchu. Może wszyscy jesteśmy ślepi na tło, w którym jesteśmy zanurzeni?

|
Co się dzieje, kiedy patrzymy na jakiś nieruchomy przedmiot? Musimy odświeżać informację docierającą do kory wzrokowej, dlatego mamy ruchy sakadyczne oczu. To właśnie mierzą okulometry: precyzyjny kierunek spojrzenia pozwala ocenić, gdzie skupia się wzrok. Mózg ocenia, gdzie można zdobyć najwięcej cennych informacji i skupia się na tych obszarach.
Ćwiczenie: zakryj jedno oko dłonią, a drugie przyciśnij palcem tak, by zatrzymać drobne ruchy gałki ocznej. Po kilkudziesięciu sekundach cały obraz świata zniknie i zostanie ciemność. Energia aktywacji kory wzrokowej bez ciągłego odświeżania rozpływa się i zanika struktura informacji dochodzących do dalszych obszarów mózgu.
Nasz słuch nie męczy nas informacjami o hałasach wewnątrz naszego ciała, nie słyszymy jak przesuwa się pożywienie w jelicie, chociaż drgania z tym związane można łatwo zmierzyć, nie zauważamy wielu odgłosów, które sami wydajemy. Istnieje rzadkie zaburzenie, w którym ludzie słyszą po przebudzeniu wiele hałasów, bo procesy filtrujące uruchamiają się powoli.
W mózgu wszystko staje się elektrycznymi impulsami, nie ma trójwymiarowej przestrzeni. W komputerze nie ma filmu ani muzyki, są tylko komórki pamięci, które po pobudzeniu wysyłają serię elektrycznych impulsów. Odpowiednio skonstruowany odtwarzacz - układ graficzny, ekran i karta dźwiękowa - potrafi te impulsy przekształcić w coś podobnego do obrazów, które widzimy i dźwięków, które słyszymy. Wystarczy odtworzyć relacje przestrzenne pomiędzy pobudzeniami pikseli ekranu by zobaczyć kształty, kolory i dźwięki. W mózgu reprezentacja jest oczywiście inna niż w komputerze, ale ogólna analogia jest słuszna. Pojawiają się wzorce aktywacji, które interpretujemy jako obrazy i inne wrażenia zmysłowe.
Jak do tego dochodzi? Dokąd biegną elektryczne impulsy z fotoreceptorów?
B17.2 Szlaki wzrokowe
Komórki zwojowe wysyłają informacje o kilkunastu różnych "ścieżkach wideo" z których tworzy się w mózgu obraz świata.
Nawet dla prostego pobudzenia małego fragmentu siatkówki impulsem świetlnym 12 komórek zwojowych wysyła odmienne "ścieżki filmowe" w głąb mózgu.
Obrazy na filmie pochodzą z neuromorficznego obwodu scalonego, który przetwarza informację w podobny sposób do siatkówki.
Każda grupa komórek filtruje specyficzne cechy obrazu: kontury, cienie, tekstury, oświetlone powierzchnie. Każdy strumień informacji jest aktywny tylko przez milisekundy. To skomplikowany system, nic dziwnego, że halucynację i zniekształcone widzenie jest dość częste.
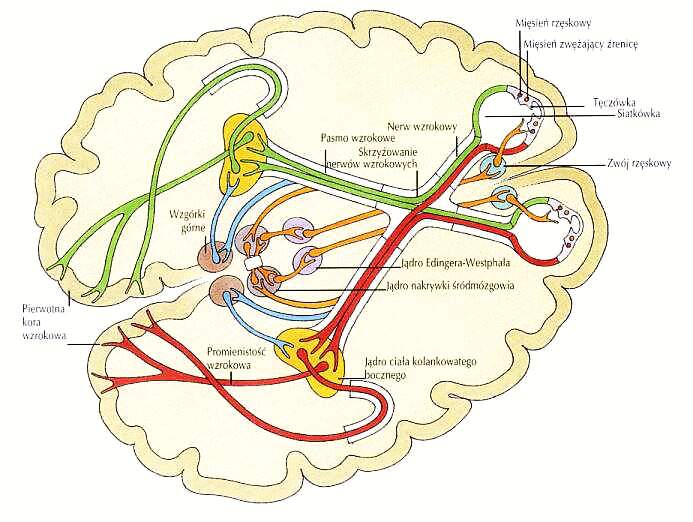
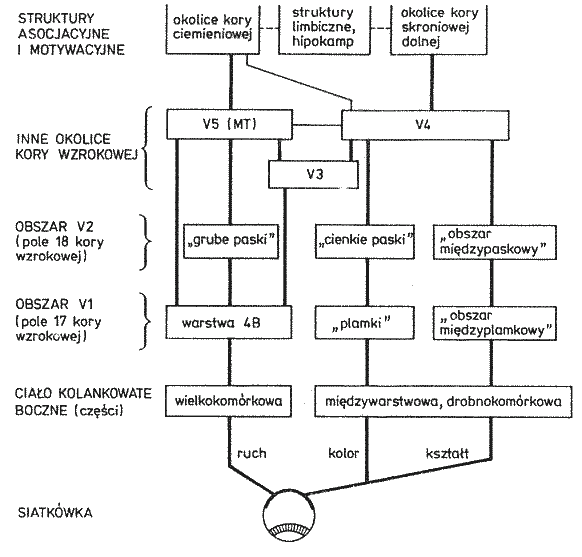
Szlaki wzrokowe: siatkówka, 6 mln czopków, 120 mln pręcików => ok. milion aksonów => ciało kolankowate boczne wzgórza => promienistość wzrokowa => obszar pierwotnej kory wzrokowej V1 => wyższe piętra układu wzrokowego => obszary kojarzeniowe i wielomodalne.
Szlaki wzrokowe.
Kora
obszaru V1, zwana jest również korą prążkowaną (białe paski na szarym tle, aksony promienistości wzrokowej kończące się w warstwie 4).
V1 zawiera komórki zorganizowane w kolumny dominacji dwuocznej i kolumny orientacyjne,
retinotopicznie (bliskie komórki reagują na bliskie sobie punkty w polu widzenia).
Proste komórki warstwy 4 reagują na paski o określonym nachyleniu, kontrastowe krawędzie, pobudzenia z jednego oka.
Znaczna część środkowego obszaru V1 reaguje na sygnały dochodzące od
okolic plamki żółtej (dołka środkowego) oka, gdzie gęstość receptorów jest największa. Sokoły i niektóre orły mają niezwykle ostry wzrok gdyż mają dwie żółte plamki.
{kind=link}
Komórki złożone w pozostałych warstwach V2-V5 reagują na sygnały z obu oczu.
Prawidłowy rozwój
układu wzrokowego wymaga odpowiedniej stymulacji w dzieciństwie, np. kora nie nauczy się interpretować różnorodnych kształtów jeśli kot chowa się w klatce tylko z pionowymi paskami.
|
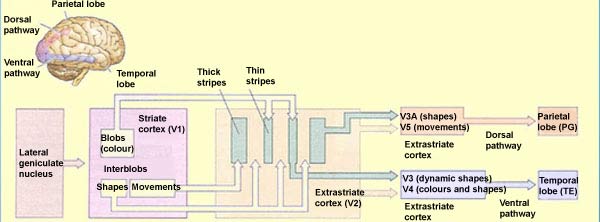
Dwa strumienie informacji wzrokowej, Ungerleider i Mishkin (1982): istnieją dwa w znacznej mierze rozdzielone szlaki przetwarzania informacji wzrokowej, biegnącej już od oka.
Wielkoziarniste komórki PA siatkówki, 3 typy stożków fotorecepcyjnych, duże pola recepcyjne, szybko przewodzące aksony, pobudzenie dla światła w szerokim paśmie.
Drobnoziarniste komórki PB, 1 lub 2 typy stożków fotorecepcyjnych, małe pola recepcyjne, wolno przewodzące aksony, rozpoznają opozycje barw.
|
Szlak wielkokomórkowy: biegnie do dwóch wielkokomórkowych warstw LGN (jest w nich ok. 100.000 komórek), charakteryzuje go niska rozdzielczość przestrzenna, wysoka wrażliwość na kontrast, szybkie przesyłanie sygnałów, bez informacji o kolorze.
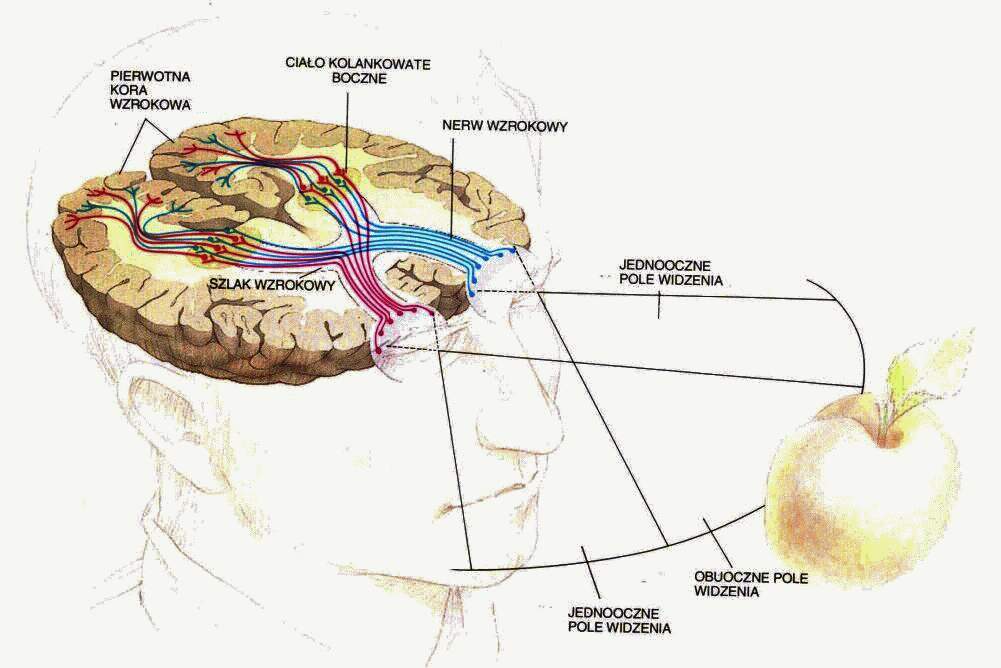
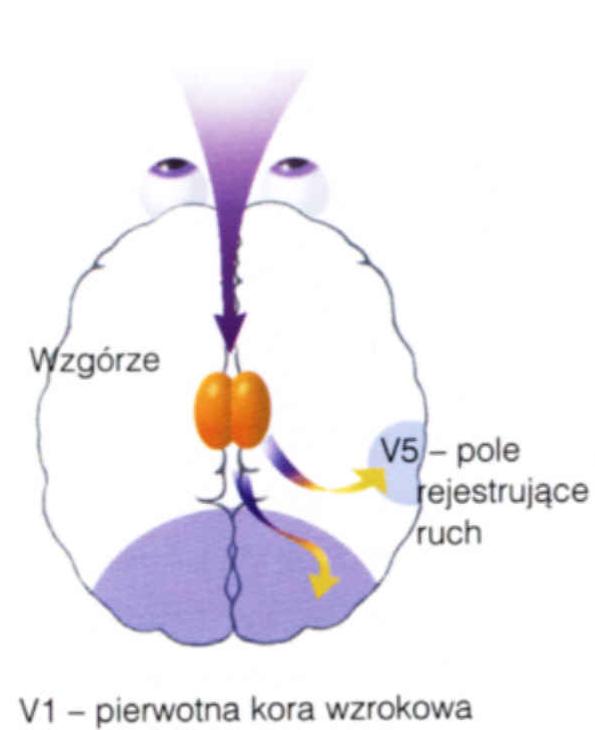
Płat potyliczny to miejsce projekcji sygnałów z nerwu wzrokowego, w którym mieści się pierwszorzędowa kora wzrokowa, poniżej bruzdy ciemieniowo-potylicznej, wyraźnie widocznej na powierzchni przyśrodkowej.
Czemu kora wzrokowa nie jest blisko oczu, tylko dokładnie po drugiej stronie głowy?
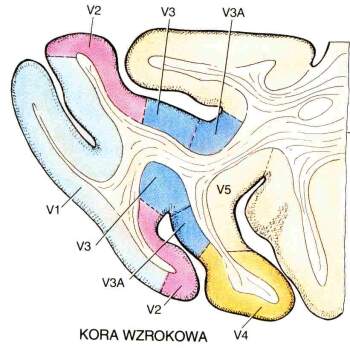
Obszary Brodmanna: 17 czyli V1, kora prążkowa, pierwotna kora wzrokowa.
Obszar 18, czyli V2, drugorzędowa okolica wzrokowa, połączona obustronnie z V1 i wysyłająca sygnały do wyższych pięter V3-V5.
Obszar 19, czyli okolice V3-V5 (strumień grzbietowy, w stronę kory ciemieniowej).
Im wyżej w hierarchii tym silniejsza jest modulacja aktywności związana z uwagą, w V1 jest ona słaba, w V4 silna.
Informacja przenoszona szlakiem wielkokomórkowym trafia przez płat potyliczny
szlakiem grzbietowym do kory ciemieniowej.
Dochodzi do warstwy 4B w V1, stąd do grubych ciemnych pasków obszaru V2, analizuje informację o ruchu obiektu.
W V1, warstwa 4B => V5, lokalizacja w polu widzenia, reakcje na ruch.
V5 pobudza płat ciemieniowy, PPC (tylna kora ciemieniowa), obszar 7 i 5; umożliwia to orientację przestrzenną, postrzeganie głębi i ruchu, połączenie z wzgórkami czworaczymi (orientacja oczu).
Szlak drobnokomórkowy ma 4 drobnoziarniste warstwy i 10 razy więcej komórek niż wielkokomórkowy w LGN.
Duża rozdzielczość przestrzenna, kolor, wolniejsze przesyłanie informacji, niska wrażliwość na kontrast.
Ta informacja trafia szlakiem brzusznym do kory dolnoskroniowej.
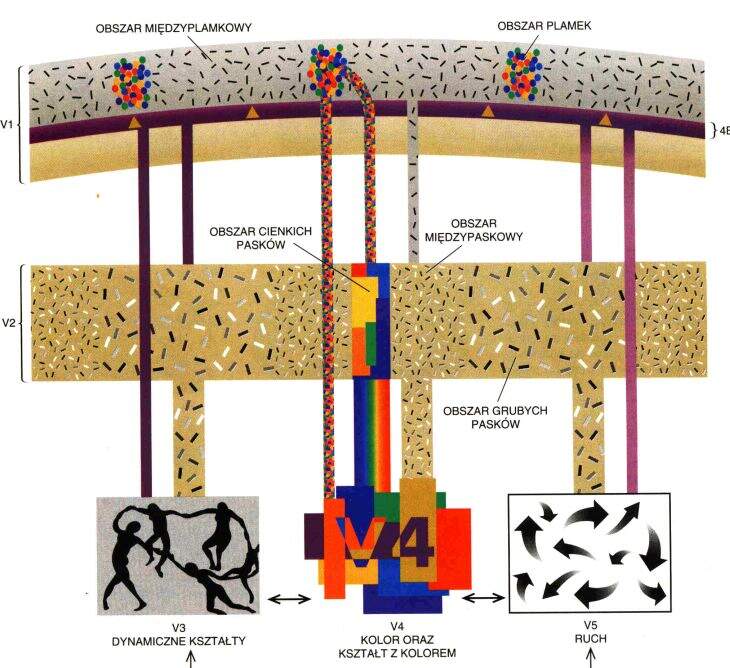
V1 => V2 obszar międzyplamkowy, reaguje na orientację linii, daje dużą ostrość widzenia, bez koloru.
V1 => V3 obszar plamkowy, reaguje na kształty, reakcja na kolor w neuronach w ciemnych prążkach V3.
V2 => V4, główny obszar analizy koloru, informacja dochodzi do kory dolnoskroniowej (IT).
Obszar IT w płacie dolnoskroniowym ma neurony reagujące na złożone obiekty.
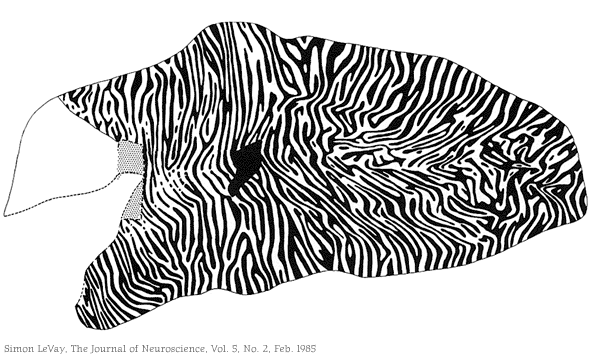{kind=link}
{kind=link}
LGN ma tylko 10-15% pobudzeń z siatkówki, pozostałe 85-90% to wsteczne projekcje z kory wzrokowej. Żeby nauczyć się widzieć trzeba powoli zgromadzić sporo informacji w głowie!
|
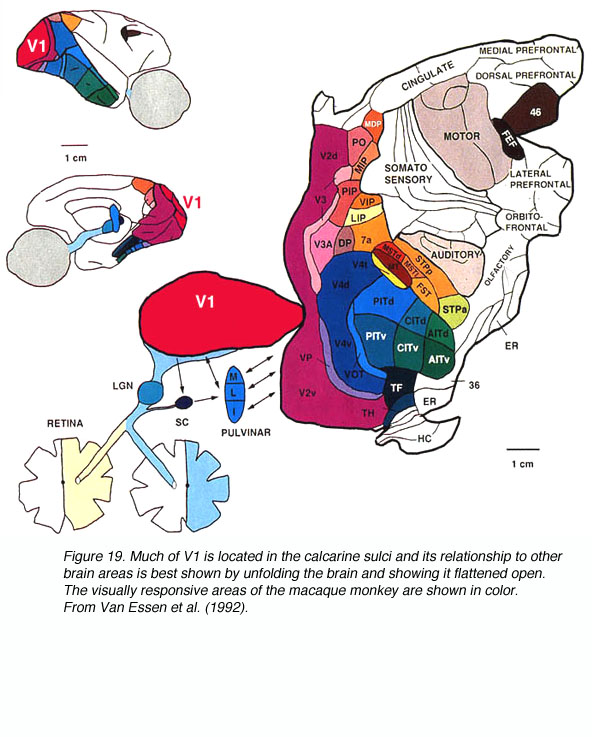
Szkic podobszarów układu wzrokowego.
Makak: dokładniejszy schemat istotnych obszarów,
oraz
schemat całości
i rozpłaszczona kora.
Rysunek podsumowujący przebieg obu
szlaków wzrokowych.
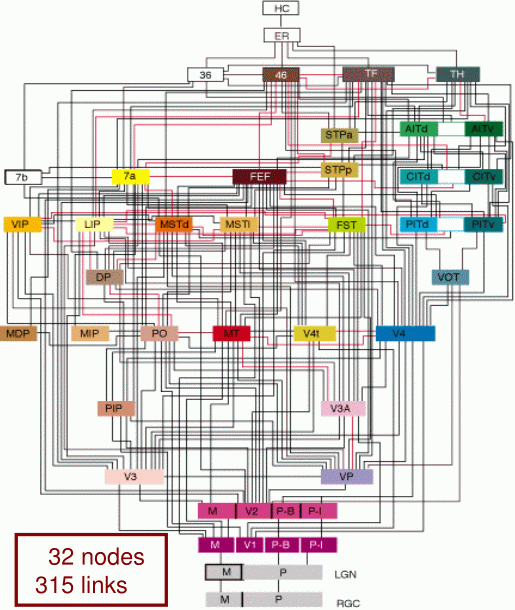
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Mamy ponad 25 obszarów kory związanych z przetwarzaniem informacji wzrokowej, zorganizowanych w hierarchiczny sposób. Dlaczego te obszary są tak liczne? Wydobywają one z sygnałów pochodzących z siatkówki niezależne od siebie informacje, porównują z wcześniej zapamiętanymi.
Położenie w przestrzeni jest niezależne od kształtu obiektu i jego wyglądu. Informacja o położeniu potrzebna jest do orientacji w stronę postrzeganego obiektu i należy ją przesłać do kory ruchowej, przyda się też do korelacji pomiędzy czuciem i wzrokiem. Postrzeganie ruchu wiąże się z położeniem w przestrzeni, stąd te obszary powinny być blisko siebie, w płacie ciemieniowym.
Wygląd potrzebny jest by rozpoznać, czym jest postrzegany przedmiot, rozpoznać zwierzę lub człowieka, skojarzyć z możliwościami działania, zaplanować to działanie, nazwać go i opowiedzieć o tym. Ta informacja powinna trafić do płata skroniowego a stamtąd do obszarów czołowych.
Stąd potrzeba różnych szlaków, grzbietowego i brzusznego.
B17.3 Teorie percepcji wzrokowej
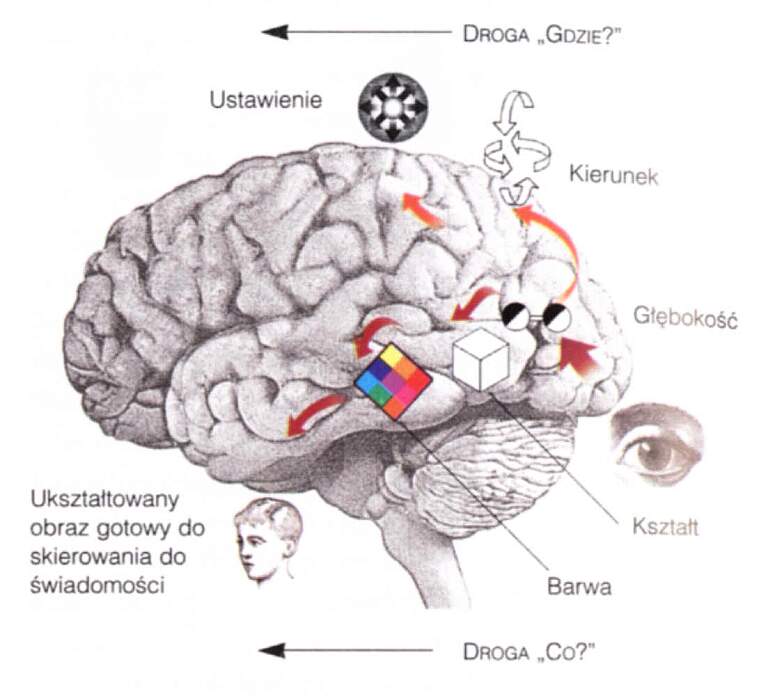
Hipoteza Ungerleidera-Mishikna:
- "co widzimy" = szlak drobnokomórkowy zmierzający do obszarów IT,
- "gdzie to jest" = szlak wielkokomórkowy, zmierzający do płata ciemieniowego.
|
Sygnały wzrokowe przez wzgórki czworacze górne i wzgórek wzrokowy sterują sakadowymi ruchami oczu. Wzgórek wzrokowy ma połączenie z korą ciemieniową, przechowująca mapę umożliwiającą orientację w przestrzeni.
Obszar zakrętu dolnoskroniowego IT ma bezpośrednie projekcje z siatkówki, pozwalając na szybkie niedokładne pobudzenie najwyższych pięter układu wzrokowego: generuje to hipotezę wstępną - do czego pasują sygnały docierające z oczu, co właściwie widzimy? Strumienie informacji zstępujące do niższych pięter pomagają w precyzyjnym rozpoznaniu: dopiero stany rezonansowe powstałe w wyniku pętli IT-V1 są uświadamiane.
Kora ciemieniowa (PC) ma wiele funkcji dotyczących przetwarzania informacji przestrzennych pozwalających na orientację. Dochodzą do niej sygnały z kory analizującej informację z kilku zmysłów: wzroku, słuchu, czucia i równowagi, co pozwala utworzyć układ odniesienia określający kierunek ruchu głowy i ciała, wyobrażenie siebie w przestrzeni. Jest to część grzbietowego szlaku przetwarzania informacji wzrokowej.
Końcowym etapem brzusznego szlaku przetwarzania informacji wzrokowej jest kora dolnoskroniowa (ITC). Neurony tam się znajdujące łączą informacje wydobytą z sygnału wzrokowego przez różne obszary układu wzrokowego, otrzymują też informację z kory ciemieniowej o położeniu obiektów. Neurony w tym obszarze reagują na złożone obrazy, utrzymują swoją aktywność pozwalając na rozpoznawanie obiektów widzianych przez krótki czas.
Analiza złożonych scen wymaga zaangażowania struktur związanych z pamięcią, umożliwiających szybkie zapamiętywanie na niezbyt długi okres czasu. Informacja z ICT trafia do kory formacji hipokampa, która kojarzy rozpoznany obiekt z kontekstem, w którym został rozpoznany, oraz tworzy mapę przestrzenną umożliwiającą nawigację w oparciu o relacje pomiędzy położeniem obserwatora i rozpoznanymi obiektami (egocentryczne i allocentryczne). Najważniejsze obszary formacji hipokampa to kora okołowęchowa (Perirhinal cortex, PRC) i
kora śródwęchowa (Entorhinal Cortex, ENC).
Kora okołowęchowa zapamiętuje relacje pomiędzy pozycjami obiektów w scenie, przydatne dla pamięci rozpoznawczej, poczucia znajomości (familiarity responses) pozwalającego na przypominanie kontekstu obserwacji.
Kora śródwęchowa jest ważnym obszarem zaangażowanym w pamięć deklaratywną. Sam hipokamp analizuje i zapamiętuje informację przestrzenną, umożliwiając jej przywołanie na podstawie wskazówek kontekstowych. Mając liczne połączenia z korą mózgu potrafi przywołać wrażenia przeżywane w trakcie zapamiętanego epizodu pobudzając odpowiednie fragmenty kory.
Częścią formacji hipokampa jest też
zakręt zębaty (dentate gyrus, DG). Jego rolą jest tworzenie wyraźnie odróżniających się od siebie wzorców aktywności, pozwalających na rozróżnienie zapamiętanych epizodów (informatyk porównał by to z
techniką haszowania).
Szybkie reakcje organizmu na niespodziewane bodźce jeszcze przed rozpoznaniem obiektu umożliwia szlak grzbietowy (kora ciemieniowa, a potem ruchowa). Postrzeganie ruchu zwierzęcia lub człowieka jest jednak związane z rozpoznawaniem charakteru ruchu. Kilka świetlnych punktów przyczepionych do rąk, nóg, głowy i tułowia, poruszających się w skoordynowany sposób jako plamy świetlne w ciemności, wystarczą by rozpoznać, że to ruch człowieka. Postrzeganie "biologicznego ruchu" zachodzi w szlaku brzusznym, w szczególności w tylnej części górnego zakrętu skroniowego (STS) (Vos i inn. 2013).
Grzbietowy szlak związany z aktywnością kory ciemieniowej analizuje bodźce wzrokowe pod kątem: "gdzie to jest w relacji do mnie i co mogę z tym zrobić", a nie tylko na pytanie ogólne "gdzie?"
Zamrożenie obszaru V5 u makaka powoduje niezdolność obszaru V2 do właściwej reakcji na docierające bodźce; nie ma hierarchii aktywacji w układzie wzrokowym, tylko współpraca każdego obszaru z innymi, sprzężenia są silne.
Świadomość wzrokowa, percepcja obiektów, zależy od pobudzenia szlaku skroniowego.
Milner i Goodale (1995): szlaki wzrokowe nie tyle określają co i gdzie, co umożliwiają działanie i percepcję.
Jest to uproszczenie, bo jest jeszcze stary szlak limbiczny, umożliwiający szybkie działanie w niebezpiecznych sytuacjach (po którym następuje fala strachu).
Potknięcie, szybkie odzyskanie równowagi i fala strachu to reakcja starego szlaku.
|
Psychofizyka próbuje powiązać fizyczne własności bodźców z subiektywnymi wrażeniami, np. intensywność światła, kontrast oświetlenia lub długość fali świetlnej z wrażeniami wzrokowymi. Zgodnie z prawem Webera-Fechnera różnice intensywności wrażenia nie wynikają z różnicy siły bodźców fizycznych, tylko z ich stosunku, zmieniając się tak jak logarytm tego stosunku. To oznacza, że dwukrotny wzrost odczuwanego kontrastu wymaga znacznie większej zmiany siły fizycznego bodźca.
B17.4 Złudzenia wzrokowe
O widzeniu można się wiele dowiedzieć badając złudzenia wzrokowe.
Widzimy to, na co jesteśmy przygotowani. By coś dostrzec musimy na tym skupić uwagę. Magicy wykorzystują to w swoich iluzjach.
Przykład:
test uwagi.
Rola kontekstu w formowaniu się obrazu jest widoczna
na obrazkach pokazanych tutaj.
Lokalny pokaz wybranych złudzeń: warto zajrzeć na stronę
Akiyoshi Kitaoka.
Uwaga! Złudzenia ruchu Kitaoki mogą wywołać dezorientację i odruchy wymiotne. Nawet
koty ulegają takim złudzeniom.

|
Złudzenia dynamiczne: odpowiednio ruszając długopisem lub prętem można wywołać złudzenie wyginania sztywnego obiektu. Po angielsku tą iluzję nazywa się rubber pencil trick.
Co się stanie, jeśli ubierzemy okulary dające żółty obraz w prawym polu widzenia, a niebieski w lewym?
Sensomotoryczna teoria widzenia mówi, że ruchy oczu są niezbędnym warunkiem by nauczyć się widzieć.
Siatkówka nie ma równomiernego rozłożenia czopków, światło różnie się odbija od powierzchni, a jednak mamy wrażenia stałości kolorów.
Nastąpi dość szybka adaptacja do widzenia przez kolorowe szkła. Już po 40 min. zdjęcie takich okularów powoduje, że w czasie ruchu oczu w prawo biała powierzchnia widziana jest jako niebieskawa (kompensując żółty kolor), a w lewo jako żółtawa (kompensując kolor niebieski). Ta zmiana postrzegania koloru zależy od kierunku i długości sakady. To pokazuje, że widzenie koloru zależy nie tylko od sygnału z receptorów ale też neuronalnych mechanizmów związanych ze sterowaniem gałką oczną.
|
Wniosek: działanie, eksploracja świata za pomocą jakiegoś zmysłu, jest konieczna do stworzenia specyficznych wrażeń z nim związanych.

|
Chociaż to niewiarygodne lewe i prawe oko ma dokładnie ten sam kolor, tzn. niebieskie i żółte oczy są szare, a czerwone jest ciemnozielone;
wystarczy je pokazać bez tła, powiększając fragment oka - wokół źrenicy widać kolor szary lub zielony.




|
Publikacje psychologiczne w latach 1950 donosiły, że ludzie mają w większości czarno-białe sny, ale w późniejszym okresie sny stały się w znacznej mierze kolorowe.
Czarno-białe sny mogły być wynikiem oglądania czarno-białej telewizji i filmów (Schwitzgebel 2002; Murzyn, 2008).
Zmiany mogą wynikać z rozpowszechnienia się kolorowej telewizji, lub szerszego używania środków farmaceutycznych.
Jednakże dane są kontrowersyjne i wygląda to tak, że wiele osób nie potrafi określić czy ma wrażenia koloru, czy nie, zapewne aktywacje ich mózgów nie przypominają dostatecznie aktywacji na jawie by można je było kategoryzować używając tych samych pojęć.
Co się stanie, gdy aktywacji ulega sam obszar IT, związany z rozpoznawaniem obiektu, a pozostałe pola wzrokowe nie są silnie pobudzone? Powinniśmy mieć wrażenie rozpoznania obiektu pomimo braku wyraźnych wrażeń kształtu czy koloru. Dowolne kombinacje są możliwe i mogą prowadzić do dziwnych wrażeń, np rozpoznania i poczucia ruchu bez koloru i kształtu.
Jeśli faza REM snu służy konsolidacji pamięci to tylko te aspekty, które są istotne powinny ulegać pobudzeniu by nie tracić niepotrzebnie energii.
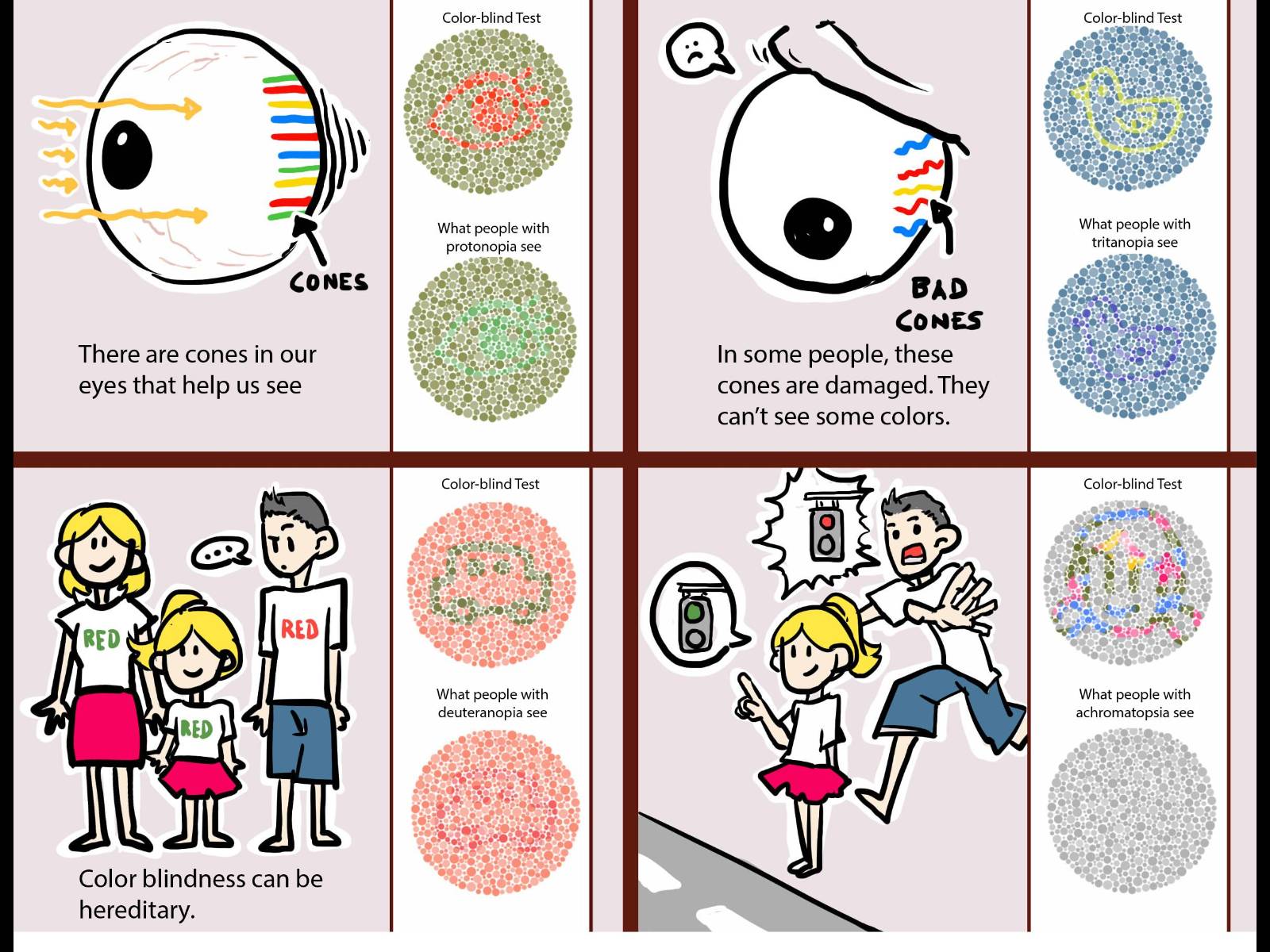
Istnieją duże indywidualne różnice w postrzeganiu kolorów;
daltonizm to skrajny przykład.
Każdy czopek może rozróżnić około 100 barw, więc trzy rodzaje czopków rozróżniają razem około 100x100x100 czyli miliona barw.
Istnieją kobiety posiadające
cztery odmienne czopki. Być może u kobiet jest to równie częste jak daltonizm, ale ponieważ zdolność widzenia dodatkowych kolorów nie jest wykorzystywana, trudno ją zauważyć. Maksimum czułości czopków może się zmienić na skutek mutacji genów na jednym z chromosomów X. Ponieważ kobiety mają dwa chromosomy X możliwe jest powstanie 4 maksimów czułości, ale u mężczyzn są chromosomy XY, więc mutacja może spowodować jedynie przesunięcie któregoś maksimum czułości, a nie powstanie dodatkowego.
{kind=link}
Jak odróżniamy halucynacje od rzeczywistości? Nie każdy je dobrze odróżnia.
Wygląda na to, że monitorowanie tego, co jest rzeczywistością zależy od aktywności i ukształtowania bruzdy czołowej okołoobręczy (paracingulate sulcus, PCS), w obszarze przyśrodkowo-przedniej kory przedczołowej (medial anterior prefrontal cortex, maPFC).
M. Buda et al, A Specific Brain Structural Basis for Individual Differences in Reality Monitoring(2011) pokazano, że indywidualne różnice w monitorowaniu swoich wewnętrznych stanów, rozróżnieniu pomiędzy percepcją a wyobrażonymi sytuacjami, mogą być związane z różnicami w budowie kory przedczołowej.
Czy niewidomi coś widzą w snach? Jeśli urodzili się niewidomi lub stracili wzrok przed 5 rokiem życia to ich sny mają inny charakter, są to głównie odczucia kinestetyczne, dotykowe czy słuchowe. Jeśli utracili wzrok w późniejszym okresie to początkowo mają wzrokowe wyobrażenia senne, ale zanikają one z upływem lat.
Badania niewidomych osób, którym
przywrócono wzrok pokazały, że potrafią one odróżnić kulę od sześcianu, ale muszą się dopiero nauczyć powiązać wrażenia wzrokowe z dotykowymi. Podejrzewał to już John Locke, który korespondował na ten temat z filozofem Williamem Molyneux, ale badania eksperymentalne
przeprowadzono dopiero w XXI wieku w ramach
"projektu Prakash".
Dzieci, które odzyskały wzrok, dostrzegają za to od razu różnice długości kresek w
iluzjach Ponzo
i Müller-Lyer
tak jak osoby widzące.
Osoby głuche od urodzenia mogą mieć kolorowe sny z komunikacją w języku migowym.
Wrażenia wzrokowe można wywołać nawet
w całkowitej ciemności poruszając swoimi kończynami. Jest to pewnego rodzaju
synestezja propriocepcji z wzrokiem.
YouTube: Visual illusions, wstęp i dyskusja, czego nas uczą złudzenia.
B17.5 Agnozje i zaburzenia wzrokowe
Kiedy wszystko działa normalnie nie zauważamy, że stoi za tym złożona maszyneria, ale kiedy się coś popsuje ...
Uszkodzenia przepływu informacji wywołują różne agnozje, zespoły neuropsychologiczne.
Uszkodzenia (udary, wypadki, zmiany neurodegeneracyjne) obszaru V2 wywołują zaburzoną percepcję kształtów.
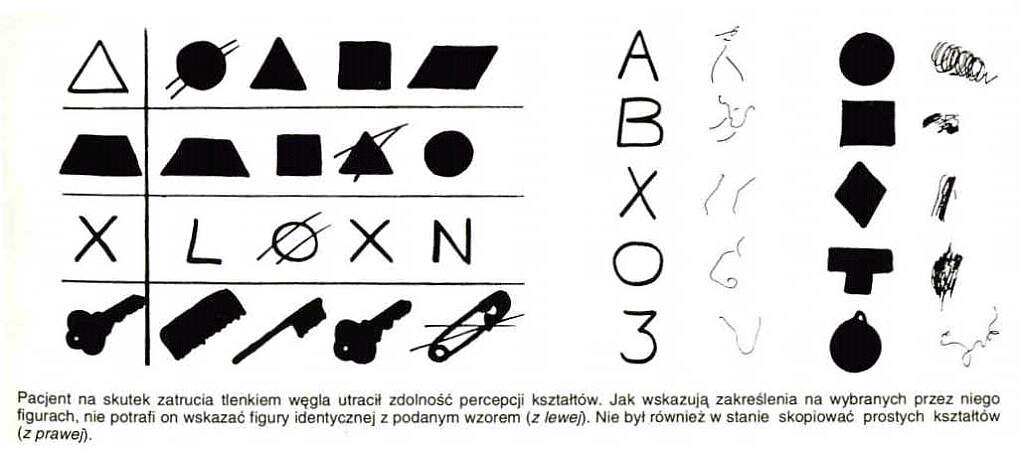
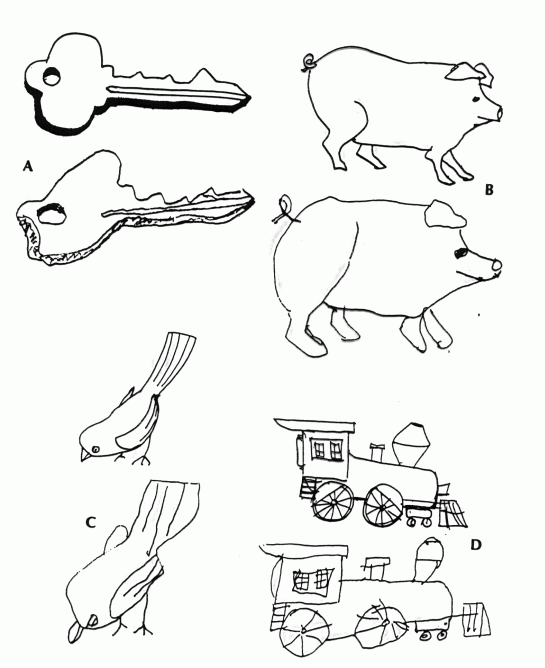
Agnozja kształtu ma wiele form, nawet proste kształty mogą być niewłaściwie rozpoznawane.
Widać to w testach selekcji lub kopiowania widzianej figury, lub pisania liter czy znaków .
Pomimo prawidłowego wykonania kopii rysunków pacjent nie ma pojęcia, co przedstawiają narysowane przedmioty.
|
Agnozja kształtu może być wybiórcza, np. bez prozopagnozji, czyli nie dotycząca twarzy. Pacjent widzi twarz ale nie poznaje warzyw, owoców i kwiatów, z których się ona składa.

|
Uszkodzenia obszaru V4 prowadzą do
mózgowej achromatopsji, ślepoty barw, czyli zaniku zdolności do widzenia kolorów.
Wrodzona achromatopsia
jest wynikiem mutacji genetycznej prowadzącej do defektu czopków w siatkówce, może być endemiczna,
np. na jednej z wysp Norwegii i Mikronezji (Sacks,
Wyspa daltonistów i wyspa sagowców) większość społeczeństwa niezdolna była do widzenia kolorów. Sacks przanalizował jak to wpłynęło na ich życie.
Anomia barw nie jest związana z percepcją ale zaburzeniami nazywania barw, są to uszkodzenia w obszarach trzeciorzędowych (zakręt kątowy).
Anomia wzrokowa może przejawiać się trudnościami z uporządkowaniem kolorów, pomimo widzenia barw - czy to jest wynik braku zrozumienia koncepcji barwy? Raczej niezdolność skojarzenia nazwy z kolorem, a więc anomia nazywania, która może występować w różnych modalnościach zmysłowych.
Jakie inne wrażenia zmysłowe większość ludzi nie potrafi precyzyjnie nazwać?

|
|
Obszar kory wzrokowej zwany V5 lub MT (Middle Temporal visual area) leży poza płatem potylicznym. Neurony w tym obszarze reagują w selektywny sposób na ruch, rozpoznając kierunek ruchu. Wystarczą do tego sygnały bezpośrednio z wzgórza, co pozwala skrócić czas reakcji na ruch. Dokładniejsze informacje dochodzą drogą przez pierwszorzędowe obszary kory wzrokowej V1, V2 i V3. Obszar V5 jest kluczowy dla postrzegania ruchu, integracji informacji na temat ruchu w całościowej percepcji, oraz kontroli ruchów gałki ocznej.
Uszkodzenia obszaru V5 prowadzą do
akinetopsji - widać wówczas tylko statyczne migawki, wrażenia przypominają widoki w świetle stroboskopowym z długimi przerwami, jakby jedna klatka filmu wyświetlana była co sekundę lub wolniej.
Krótki film ilustruje ten rodzaj agnozji, zwany również "ślepotą ruchu".
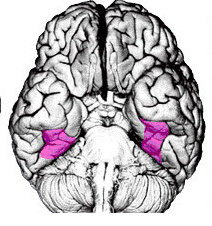
Obszar Brodmanna 37, czyli potyliczno-ciemieniowy zakręt wrzecionowaty
(fusiform gyrus) jest dużym obszarem brzusznej części kory, rozciągającym się od płatów potylicznych do skroniowych. Część nazywana wrzecionowatym obszarem twarzy
(FFA, fusiform face area) specjalizuje się w rozpoznawaniu twarzy.
Uszkodzenia obszaru FFA powodują
prozopagnozję, niezdolność do rozpoznawania twarzy; w tym obszarze ponad 90% komórek reaguje tylko na twarze. Wszystkie twarze wydają się wówczas podobne (jak nam np. twarze Chińczyków), ale można nawet nie odróżniać własnej twarzy od innych.
Czasami pomimo braku rozpoznania da się zaobserwować reakcje emocjonalne na poziomie podkorowym, np. reakcje GSR (czyli zmianę oporności skóry na skutek pocenia w wyniku takich reakcji).
Zachowana jest zdolność do rozpoznawania zwierząt (np. indywidualnych owiec w przypadku pasterza).
Normalnie układ wzrokowy się uczy, mózg przystosowuje się do nowych bodźców; w prozopagnozji nie potrafi się już nauczyć gdyż kora w tym się specjalizująca uległa uszkodzeniu.
Czy rozpoznawanie twarzy jest wrodzone czy wyuczone? Małpy i niemowlęta reagują na twarze, ale też reagują na proste twarzo-podobne rysunki, co wskazuje raczej na ogólne preferencje mózgu do rozpoznawania kształtu głowy, położenia nosa, oczu i ust. Pasterze na Nowej Zelandii rozpoznają lepiej pyski owiec niż ludzkie twarze, co wskazuje na procesy uczenia się.
Są przypadki braku zdolności rozpoznawania od urodzenia. Prozopagnozja apercepcyjna to właśnie brak zdolności do identyfikacji twarzy indywidualnych osób. Prozopagnozja asocjacyjna wynika z braku możliwości połączenia rozpoznania z informacjami na temat danej osoby. Jedna z częstych form takiej prozopagnozji to trudności z przypominaniem sobie imienia czy nazwiska rozpoznanej osoby.
Przypomina to anomię , czyli afazję nominalną.
Jak widać prozopagnozja może występować w różnej formie i stopniu.
Dlaczego u wszystkich ludzi akurat w tym miejscu znajduje się obszar reagujący na twarze?
Zapewne znajdujące się tam sieci neuronowe mają połączenia do tych obszarów wzrokowych, które wyodrębniają specyficzne cechy przydatne do rozpoznawania twarzy. Nie mamy jeszcze szczegółowych modeli wyjaśniających takie procesy. Największe nadzieje wiążą się z teoretycznymi modelami samoorganizacji sieci neuronowych mózgu. Jakie genetycznie uwarunkowane struktury potrzebne są by rozwinąć taką zdolność i jakie bodźce są do tego konieczne? To nadal otwarte pytania.

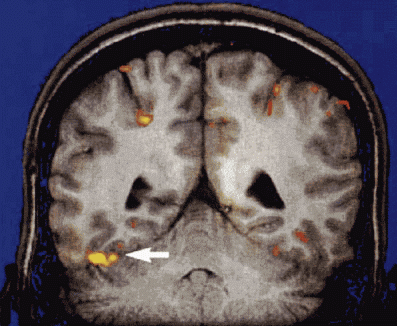
Analiza fMRI procesu rozpoznawania twarzy. Widać wyraźną lokalizację aktywności w obszarze FFA. |
|
Wrażenia wzrokowe to rezultat działania wyższych pięter układu wzrokowego, w płatach skroniowych.
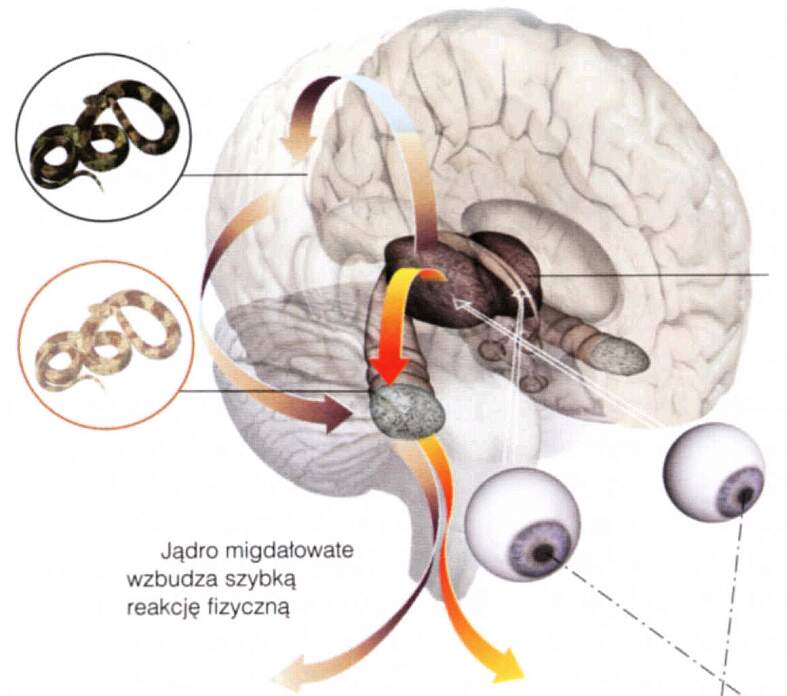
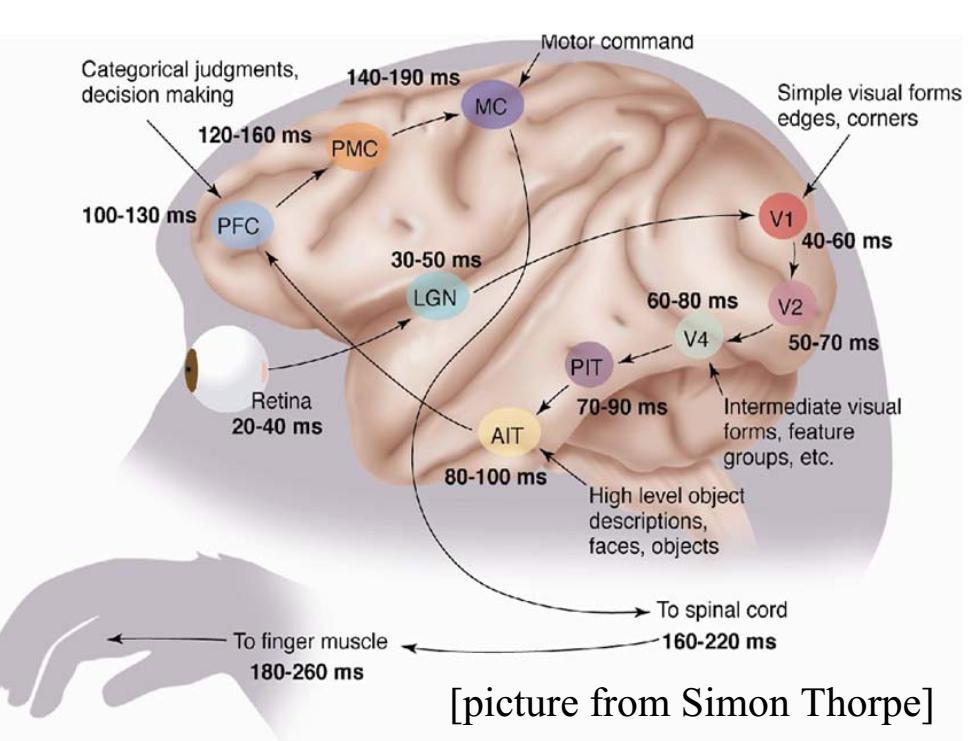
Droga świadomego działania na wzrokowy bodziec od oka do ręki przebiega przez wzgórze (LGN, ciało kolankowate boczne), korę potyliczną (V1, V2, V4), dolny zakręt skroniowy (PIT, AIT), korę przedczołową (PFC), korę ruchową (PMC, MC), nerwy w kręgosłupie, do mięśni ręki. Zajmuje to około 180-260 ms. Na rysunku poniżej nie zaznaczono szybszej drogi prowadzącej do automatycznych, instynktownych reakcji. Prezentacja bodźców na peryferiach pola widzenia wywołujących strach prowadzi do aktywacji ciała migdałowatego w czasie krótszym niż 80 ms, umożliwiając reakcje ruchowe w ciągu 1/10 sekundy.
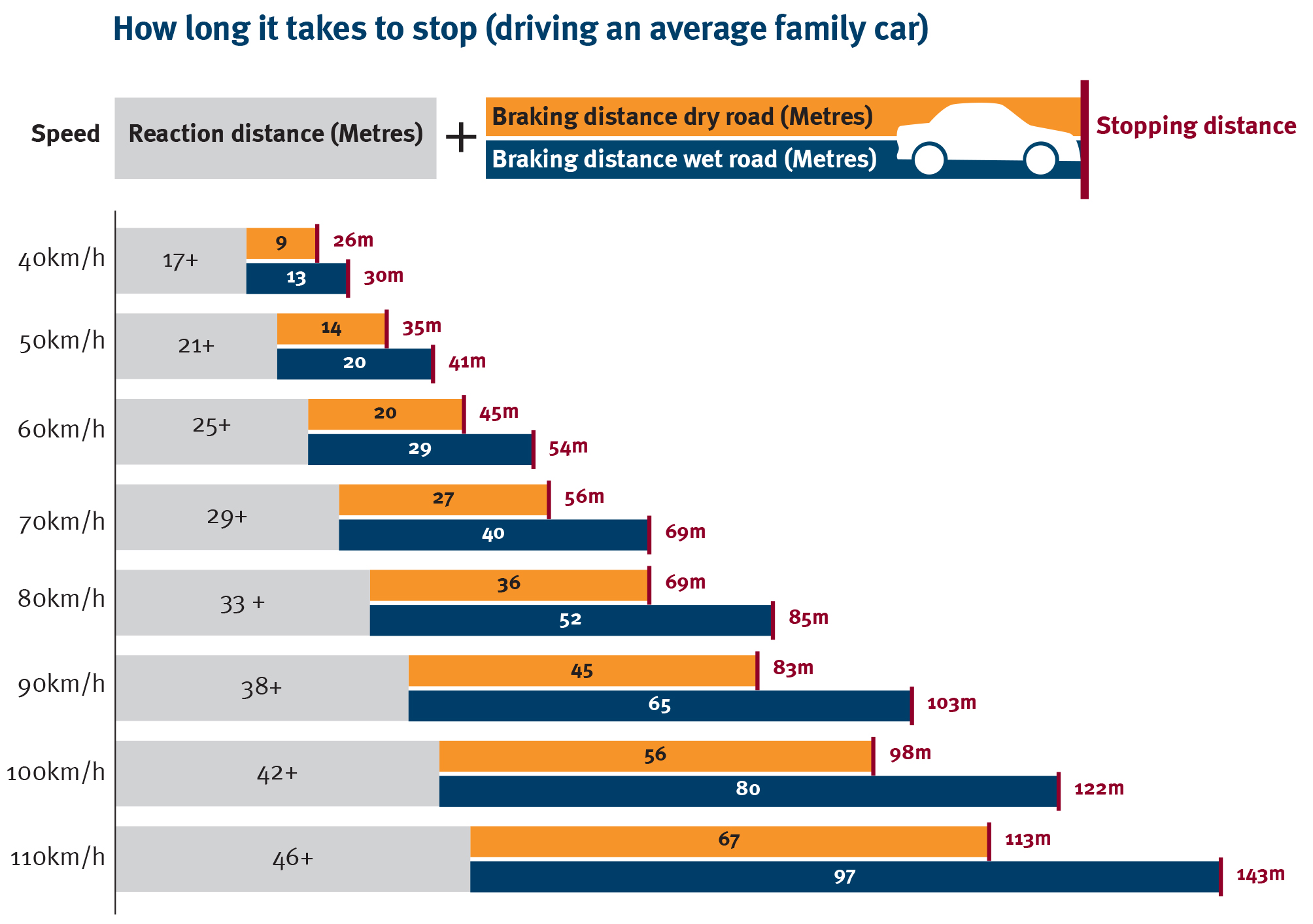
Ciekawostka: droga hamowania samochodu zależy od czasu reakcji i czynników fizycznych.
Infografika pokazuje
typowe czasy, ale czasy reakcji są tu mniej znaczące niż czynniki fizyczne: hamulce, opony, masa samochodu, stan nawierzchni.
{kind=link}
|
Pobudzenia siatkówki docierające przez nerw wzrokowy odpowiedzialne są za niewielką część aktywności powyżej V1 (około 10%). We śnie lub pod wpływem halucynogenów możemy mieć wyraźne wrażenia wzrokowe bez pobudzenia siatkówki.
Świat widziany to głównie wyobrażenia, wynikające z zapamiętanych wcześniej bodźców, oraz wewnętrznej dynamiki procesów w mózgu.
W jaki sposób tworzy się spójne wrażenie z aktywności różnych obszarów, obrazu rozbitego na różne elementy (kształt, kolor, ruch)? Powstaje problem spójności wrażeń wzrokowych i innych zmysłów
(binding problem).
Czym różnią się od siebie zintegrowane wrażenia, które możemy analizować zwracając uwagę na różne modalności zmysłowe? Są tu różne teorie, ale jest to nadal kontrowersyjne zagadnienie.
Dolny płacik ciemieniowy
(IPL, inferior parietal lobule)
zawiera trzeciorzędowe obszary kory (zakręt kątowy, zakręt nadbrzeżny, obszar Wernickiego) integrujące informacje wzrokowe, czuciowe i słuchowe. Jest więc obszarem bardzo istotnym dla rozpoznawania złożonych perceptów.
Uszkodzenia szlaku prowadzącego do zakrętu kątowego i dolnego zakrętu skroniowego (AIT, PIT) prowadzi do
agnozji wzrokowej, czyli niezdolności do nadania sensu temu co się widzi. W zależności od miejsca i rodzaju uszkodzenia niemożliwe jest wówczas świadome rozpoznanie przedmiotów przy zachowanej zdolności do działania, np. uchwycenia przedmiotu czy manipulacji nim (ślepota psychiczna).
Zaburzenie dotyczyć może zdolności rozpoznawania obiektów należących do określonej kategorii, np. samochodów, krzeseł, zwierząt czy palców ręki.
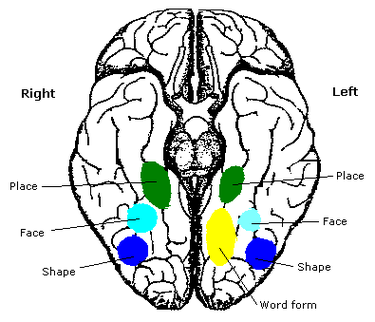
Analiza aktywności mózgu przy oglądaniu złożonych scen pokazała zwiększoną aktywność trzech regionów selektywnie kodujących informację o krajobrazie: kory przy hipokampie (parahippocampal place area, PPA), retrosplenial cortex, RSC - to kora pomiędzy formacją hipokampa i płatem ciemieniowym, za tylną częścią ciała modzelowatego (BA 28 i 29), oraz obszaru na bocznej powierzchni płata potylicznego, poprzecznej bruzdy potylicznej "occipital place area, OPA,lub transverse occipital sulcus, TOS). To obszary reprezentujące relacje przestrzenne, uwzględniając położenie powierzchni w 3 wymiarach.
Symultanagnozja (agnozja symultatywna): postrzeganie pojedynczych aspektów, ale nie całości.
Np. można widzieć poszczególne kształty, ale nie rozumieć znaczenia całości.
Widzenie całości to złożony problem, wymagający koordynacji działania kory wzrokowej, ruchów oczu, skupiania uwagi, kojarzenia informacji, spójności elementów wrażeń.
Uszkodzenie pierwotnej kory wzrokowej (np. niedokrwienie) może prowadzić do pojawienia się czarnej dziury w polu widzenia
(mroczek, skotoma), a w rozległej formie jest to
ślepota korowa.
Ubytki w polu widzenia są często dopełniane interpolowanymi danymi.
Badani są przekonani, że widzą całość (podobnie jak nie widzimy obszaru plamki ślepej), bo nauczyli się bezwiednie poruszać oczami i głową tak, by informacja wzrokowa w brakującym polu została wypełniona. Dopiero testy wzroku uświadamiają im, że nie widzą pewnych obszarów. To kolejny przykład jak silnie układ ruchowy wpływa na naszą percepcje. Więcej takich przypadków omówimy w następnym wykładzie, bo wiążą się z dotykiem i ruchem.
B17.6 Wyobraźnia wzrokowa
Wrażenia wzrokowe (świadomość widzenia) pojawiają się dzięki aktywnej eksploracji, szukania informacji i rozróżnianiu stanów układu wzrokowego na poziomie skojarzeniowym, można je uznać za wewnętrzny komentarz.
Uszkodzenie kory lub promienistości wzrokowej prowadzi do utraty wzroku, chociaż informacje z siatkówki trafiają do różnych części mózgu.
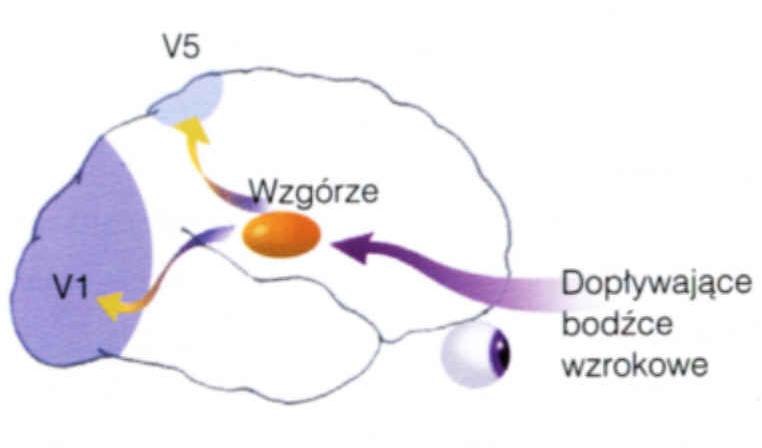
Ślepowidzenie (blindsight)
lub ślepowzrok to szczątkowe widzenie bez wrażeń wzrokowych, które może się pojawić przy takich uszkodzeniach.
Zaobserwowano je początkowo u małp, a później u ludzi (przykład: film na którym
niewidomy obchodzący przeszkody).
Zachowana jest częściowa zdolność do lokalizacji miejsca, ruchu, kształtu a nawet koloru, chociaż badani "nic nie widzą", tylko zgadują.
Informacja dociera przez wzgórze (LGN) i wzgórek wzrokowy do wyższych pięter układu wzrokowego i płata ciemieniowego.
Ilustruje to ten film.
W miarę treningu pacjenci nabierają wprawy w "wyczuwaniu" widoku, chociaż nie przypomina to wrażeń wzrokowych.
|
Ślepowidzenie dostarcza innych wrażeń, które trzeba się nauczyć interpretować. Nie są to normalne wrażenia wzrokowe, bo nie ma rozróżnienia pomiędzy stanami kory wzrokowej, które są konieczne by je porównać z zapamiętanymi stanami.
W rzadkich przypadkach takie osoby reagują emocjonalnie na wyrazy twarzy osoby z nimi rozmawiające, na ile jest to jednak reakcja wzrokowa a na ile gra tu rolę słuch? Oczywiście z ciała kolankowatego bocznego są projekcje do układu limbicznego i stąd jakaś analiza emocji powinna być możliwa, w końcu większość reakcji związanych z empatią nie jest świadoma.
Ślepowidzenie afektywne
to szczątkowa zdolność do reakcji na bodźce o zabarwieniu emocjonalnym, pomimo braku świadomości wzrokowej.
"Widzenie" u niewidomych wywołać można przez pobudzanie sygnałem z kamery skóry na plecach lub pobudzenia na języku, człowiek szybko uczy się właściwej interpretacji sygnałów, a ponieważ pozwalają one na podobną interakcję ze światem jak wzrok wrażenia są podobne. Trwają próby pobudzania bezpośrednio obszaru kory wzrokowej V1, ale wrażenia w ten sposób wywoływane są mało precyzyjne. Prawdopodobnie konieczna jest stymulacja za pomocą bardzo wielu elektrod, ale jest to technicznie trudne (projekty takiej stymulacji są szumnie zapowiadane, ale na początku 2022 roku nie było jeszcze konkretnych rezultatów).
|
Takie sygnały odczuwane są jako widzenie w sensie analizy relacji przestrzennych, chociaż wszystkie sygnały mają tą samą postać pobudzeń neuronów.
Wniosek: modalność zmysłowa w mózgu związana jest ze sposobem analizy informacji dopływającej ze zmysłów.
Zaprzeczanie ślepocie (zespół Antona)
Pomimo ślepoty badani mają wrażenia wzrokowe, odmawiają nauki z niewidomymi, usiłują sami chodzić chociaż ciągle obijają się o przedmioty i przewracają.
Mają tendencje do konfabulacji, np. opisu widzianej osoby. Lekceważą przy tym niezgodności opisu próbując je wyjaśnić racjonalizując, np. "jest noc, słabe światło, zagracony pokój".
Jest wiele innych zaburzeń, wynikających z uszkodzeń płatów potylicznych, np. palinopsja (perseweracja wzrokowa), widzenie lub wielokrotne pojawianie się obrazu po ustaniu wywołującego go bodźca.
Zaburzenie konwersyjne, nazywane wcześniej ślepotą histeryczną, to brak wrażeń wzrokowych, pomimo działającej kory wzrokowej.
Jest to jedno z wielu zaburzeń dysocjacyjnych, czyli pozornych problemów pojawiających się bez fizycznej przyczyny.
Wykrywana za pomocą odruchu obronnego, odruchu okoruchowego, badań EEG.
Zespół Charlesa Bonneta!
Proste halucynacje geometryczne pobudzają głównie V1, a twarze zakręt wrzecionowaty (FFG).
Zdeformowane twarze szczególnie pobudzają fragment FFG związany z obrazem oczu i ust (zębów); są też halucynacje związane z kreskówkami.
Halucynacje w tym zespole są raczej na niskim poziomie, wydają się jak film, zdarzają sie tylko ludziom niedowidzącym lub niewidomym, nie jest to podobne do snów bo te halucynacje nie mają jakiegoś związku z przeżyciami danej osoby.
Zespół Charlesa Bonneta opisany został w książkach Ramachandrana (Phantoms in the Brain) i Chandry (Sacred Games).
Stymulacja zakrętu IT wywołuje halucynacje wzrokowe.
Ogólnie o halucynacjach, nie tylko wzrokowych poczytać można w
tym artykule Wiki.
Czemu ludzie widzą duchy tylko w ciemności?
Układ wzrokowy dostarcza informacje pozwalając na poprawną identyfikację tylko przy dobrym oświetleniu, korelacja aktywności kory wzrokowej V1 z sygnałami dochodzącymi z siatkówki jest na poziomie 10%, więc przy ciemniejszym jeszcze słabsze, nic dziwnego, że pojawiają się halucynacje mające mało wspólnego z tym, co widzimy w pełnym oświetleniu.
Pareidolia dotyczy najczęściej twarzy czy ludzkich postaci. W przypadkowych bodźcach, zwłaszcza zmiennych jak płomień ogniska czy chmury, możemy się doszukać jakichś kształtów.
W szumie możemy też usłyszeć różne głosy. Potrzebne jest do tego pobudzenie ukła du wzrokowego lub słuchowego, który próbuje nadać bodźcom jakąś zrozumiałą interpretację.
W chaosie widzimy znajome kształty czy słyszymy głosy, bo mózg chaotycznie pobudzany ma skłonność do utworzenia jakichś spójnych ścieżek pobudzeń, interpretowanych jako rzeczywiste wrażenia; widać tu interakcję (zmysły<=>ośrodki skojarzeniowe) zarówno strumieni informacji wstępujących jak i zstępujących (por. np.
Bruce Hood, SuperSence, 2009).
Rezonans stochastyczny w układzie wzrokowym pokazuje, jak słabe sygnały mogą dzięki szumowi wywołać wrażenia wzrokowe. Szum pełni wiele funkcji w układzie nerwowym, ale czasami wywołuje halucynacje różnych zmysłów.

|
Apofenia to ogólna tendencja do doszukiwania się sensu w przypadkowych obserwacjach, nie tylko obrazów (np. twarzy w chmurach lub na zdjęciu Marsa). U osób podatnych na apofenię stwierdzono w teście plam atramentowych Rorschacha, że obszar związany z rozpoznawaniem twarzy uaktywnia im się silniej niż pozostałym badanym, dlatego cierpią na pareidolię.

|
Nawet robot może mieć takie złudzenia ... percepcja oparta jest na wyszukiwaniu w pamięci hipotez interpretacyjnych najlepiej pasujących do bodźców i w niejednoznacznych przypadkach prowadzi często do halucynacji.
Czy niewidomy może malować? Zmysł dotyku daje podobne informacje, pozwalając tworzyć wyobrażenie relacji przestrzennych, dlatego używany jest jako substytut wzroku u osób niewidomych.
Esref Armagan z Turcji, niewidomy od urodzenia, maluje używając kolorów i stosując perspektywę, np. malując oktagonalny budynek baptysterium we Florencji.
Jego kora wzrokowa reaguje na relacje przestrzenne badane przez dotyk, używane kolory często są przypadkowe, a w innych przypadkach są to wyuczone kolory znanych obiektów, np. drzewa zielone, artysta wie gdzie farby o różnych kolorach są na jego palecie (Amir Ahmed i inn, 2008).
Wyobraźnia wzrokowa związana jest prawdopodobnie z możliwościami odtworzenia w korze wzrokowej podobnych pobudzeń jak w czasie aktualnego doświadczenia. Wymaga to dostatecznie silnego pobudzenia niższych obszarów przez wyższe obszary układu wzrokowego (podobnie jest w innych układach zmysłowych).
Dla skomplikowanych obiektów potrzebna jest przestrzeń neuronalna, w której tworzy się plan działania; bierze w tym udział kora ciemieniowa.
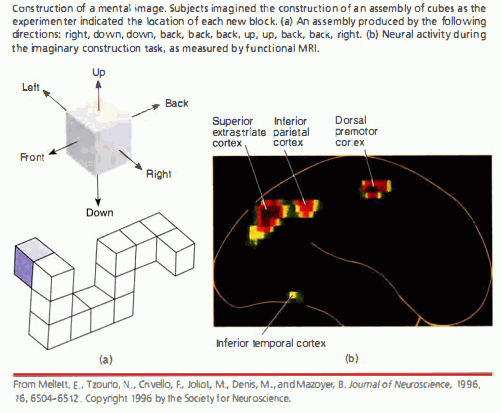
|
Istnieją silne indywidualne różnice w zdolności do wyobrażenia sobie i np. narysowania czegoś z pamięci (czy zaśpiewania lub zagrania melodii z pamięci).
Te różnice mierzy test żywości wyobraźni wzrokowej.
B17.7 Mechanizm widzenia
Na czym więc polega widzenie? Mechanizm ten z grubsza wygląda tak:
- Na poziomie siatkówki dokonuje się wstępne przetwarzanie sygnałów, lokalnych kontrastów (komórki ON/OFF), a informacje przesyłane są dwiema drogami do LGN i pierwotnej kory wzrokowej V1.
- Z LGN do obszarów dolnoskroniowych, przygotowując ten obszar do właściwej analizy i ograniczając możliwe interpretacje informacji wzrokowej.
- W korze V1 i V2 wyspecjalizowane kolumny korowe dokonują dalszej prostej analizy obrazu przy zachowaniu projekcji retinotopicznej i małych pól recepcyjnych: wydobywane są informacje o nachyleniu krawędzi, kolorze i ruchu w polu widzenia.
- Te informacje łączone są w obszarze V3 tworząc dynamiczne kształty, oraz V4 łącząc kolory z kształtami.
- Komórki leżące blisko siebie grupują się w złożone struktury; to grupowanie percepcyjne opisane zostało na ogólnym poziomie w psychologii postaci (gestaltu).
- W obszarze MT/V5 i tylnej korze ciemieniowej tworzy się mapa pozwalająca na umiejscowienie w przestrzeni oglądanego obiektu (szlak grzbietowy).
- Kora skroniowa dolna uzgadnia interpretację sygnałów z obszaru V3 i V4, skupiając się głównie na sygnałach dochodzących od obszaru na który wskazuje silne pobudzenie w obszarze V5 (skupianie uwagi, pośredniczy w tym tylna część bruzdy skroniowej górnej STS).
- Informacja o rozpoznanym obiekcie jest analizowana przez płaty czołowe, kontrola uwagi i ruchy sakadyczne oczu pomagają dostarczyć informacje pozwalającą na jednoznaczną interpretację postrzeganego obiektu.
Widzenie wymaga więc powstania sprzężenia pomiędzy najniższymi (V1) i najwyższymi (IT, V5) obszarami układu wzrokowego. Można to bezpośrednio stwierdzić hamując aktywność STS za pomocą rTMS, krótkich impulsów pola magnetycznego (Alvaro Pascual-Leone, Silvanto, Batteli i inn.); trzeba wiedzieć w którym momencie zastosować impuls, trzeba w tym celu monitorować sygnały EEG.

widać na zmianę niebieski i czerwony napis. |
Jest szereg figur, które postrzegamy w różny sposób chociaż obraz się nie zmienia. Najstarszym przykładem jest
sześcian Neckera - raz widzimy z przodu ścianę z lewej a raz z prawej strony. Innym ciekawym zjawiskiem często używanym w badaniach jest
rywalizacja obuoczna.
Kiedy jedno oko widzi obraz, który jest całkiem inny niż drugie mózg próbuje je scalić, ale skoro nie jest to możliwe tworzy na zmianę raz jeden obraz a raz drugi - wynika to z konkurencji pomiędzy grupami neuronów, które próbują się maksymalnie zsynchronizować. Nazywamy to mechanizmem "zwycięzca bierze wszystko"
(Winner Takes All (WTA)).
Zwykle używa się okularów wirtualnej rzeczywistości, ale można też zobaczyć ten efekt za pomocą okularów polaryzacyjnych 3D z kolorowymi szkłami.
W eksperymentach fMRI z rywalizacją obuoczną można sprawdzić, czy świadomość wzrokowa zależy od aktywności kory V1 - okazuje się, że nie zależy. Świadoma uwaga wpływa natomiast na stan kory V1 (Watanabe i inn 2011).
To, co widzimy, nie powinno zależeć od drobnych zmian, bo chcemy rozpoznawać przedmioty pomimo spadających kropli deszczu, migotania odblasków, cieni, przy słabej widoczności o półmroku czy we mgle. Okienko ciągłości (continuity field) jest dość długie, nawet do 15 sekund, system wzrokowy poświęca dokładność na rzecz stabilności percepcji (Fischer i Whitney, 2014).
W okienku ciągłości wiele podobnych do siebie obiektów wydaje się bardziej podobnych niż jest w rzeczywistości. Rozpoznajemy twarze, osoby czy przedmioty, nie zwracając uwagi na zmianę szczegółów.
Wynikiem tego jest ślepota zmian
(Change Blindness) - nie zauważamy powolnych zmian w obrazie, jeśli rozpoznawane najważniejsze obiekty są stałe, ani szybkich zmian, jeśli coś odwróci naszą uwagę. Są dwie główne wersje zmian, które trudno nam dostrzec. Jedna związana jest z powolnymi, ciągłymi zmianami,
w ciągu minuty
może się zmienić prawie wszystko ale większość ludzi tego nie dostrzega.
Druga możliwość to odwrócenie uwagi:
słynny goryl, wędrujący przez boisko, podmiana recepcjonisty, który na chwilę się schyla a klient się odwraca, zmiana sceny filmu powoduje, że nie pamiętamy szczegółów, które były w poprzedniej. Iluzją jest nasze przekonanie, że wszystko widzieliśmy i pamiętamy. To powoduje, że naoczni świadkowie są mało wiarygodni.
Książka C. Chabris i D. Simons,
The Invisible Gorilla: How Our Intuitions Deceive Us. Harmony, 2011, pokazuje wiele przykładów, jak złudne jest nasze przekonanie, że wiemy co się naprawdę dzieje.
Ma to
praktyczne implikacje przy ocenie wiarygodności naocznych świadków, rozpoznawaniu zmian w ruchu drogowym, czy wyszukiwaniu informacji w czasie monitoringu przy użyciu wielu ekranów.
Dotyczy to nie tylko wzroku, ale mamy też
ślepotę zmian akustycznych, dotykowych czy zapachowych.
Podsumowując: wiesz, co widzisz, i widzisz, co wiesz ...
Czy reakcja na niektóre bodźce jest wrodzona? Widząc ruch patyka, na który przypadkowo staniemy, możemy się wystraszyć i odskoczyć.
Nie na specyficzne kształty, ale może pewne cechy wydobyte z sygnału wzrokowego i słuchowego mogą być interpretowane jako niebezpieczeństwo: ruch wężowy, wielkość, kształt, oczy i zęby dużych kotowatych drapieżników, wilków lub niedźwiedzi.
Stabilne kombinacje niektórych cech mogą być wykrywane przez specyficzne filtry jeszcze przed dotarciem informacji do kory, na poziomie jąder wzgórza i ciała migdałowatego, pozwalając na mobilizację organizmu zanim jeszcze obiekt zostanie rozpoznany.
Ciekawostka:
jak psy widzą kolory?
|
Zadanie:
|
Przykładowe pytania by sprawdzić swoje wiadomości:
- Wymień sprzeczne wymagania jakie natura stawia widzeniu.
- Komórki siatkówki.
- Jak wygląda szlak wzrokowy od oka do kory? Uwzględnij podział na szlak wielkokomórkowy i drobnokomórkowy.
- Dlaczego istnieją dwa szlaki, wielkokomórkowy i drobnokomórkowy?
- Jakie obszary składają się na brzuszny szlak wzrokowy i jakie są jego funkcje?
- Jakie obszary składają się na grzbietowy szlak wzrokowy i jakie są jego funkcje?
- Czemu nie widzimy obrazu odwrotnie?
- Czemu ludzie widzą duchy tylko w ciemności?
- W procesie widzenia jaki jest wpływ informacji z siatkówki a jaki z reszty mózgu na stan kory wzrokowej?
- Hipoteza Ungerleidera-Mishikna.
- Hipoteza Milner-Goodale.
- Co to jest prozopagnozja i jakie uszkodzenia mózgu ją powodują.
- Na czym polega i z czym się wiążę agnozja kształtu? Symultagnozja?
- Co to jest ślepota korowa?
- Na czym polega ślepowidzenie i dlaczego jest możliwe?
- Czym różni się achromatopsja od anomii barw?
- Jak przejawia się zespół Charlesa Bonneta? Komu się przydarza i z czego wynika?
- Czy niewidzialny człowiek sam mógłby widzieć? Czy mógłby słyszeć?
Programy stacji Planete+HD warte są obejrzenia:
Film ilustrujący ewolucję ciała człowieka:
Historia ewolucji ludzkiego ciała.
Seria
Nasze niesamowite zmysły pokazuje wiele ciekawych informacji o działaniu zmysłów.
Niewidzialny dom, czyli co widzę jak ma zamknięte oczy?
BBC Earth w 3 odcinku serię "Dziewięć najważniejszych miesięcy", pokazuje Pingelap, wyspę na której jest 10% osób z achromatopsją.
Visual illusions, wstęp i dyskusja, czego nas uczą złudzenia.
Ciekawe strony z iluzjami wzrokowymi.
- Iluzje wzrokowe.
- Iluzje roku.
- Kwadratura koła.
- Test kolorów.
- Cool optical illusions *YouTube, 6 min 30 s).
- Color constancy illusions.
- Świetna seria "Pułapki umysłu" pokazywana przez National Geographic TV ilustruje liczne złudzenia percepcji.
- Superhuman color vision! Filtry, które pozwalają rozróżniać metamery. Jaki kolor ma ta sukienka? Jaki ubranie Nike?
Literatura:
Dobre książki o widzeniu:
- Jan Młodkowski, Aktywność wizualna człowieka Wyd. Naukowe PWN SA, Warszawa 1998
- Piotr Francuz, IMAGIA. W kierunku neurokognitywnej teorii obrazu. Wyd. KUL, Lublin 2013.
- Jaśkowski, P. (2009). Neuronauka poznawcza. Jak mózg tworzy umysł. Vizja Press. Rozdz. 5. Jak mózg steruje działaniem.
- Kolb, B., Whishaw, I. Q., & Teskey, G. C. (2019). An Introduction to Brain and Behavior. Worth Publishers.
- Carter, R., & Parker, S. (2019). The Human Brain Book. DK Publishing.
- Gary Matthews, Neurobiologia. Wyd. Lekarskie PZWL 2000, rozdział 16 i 17.
- 10 blind painters.
Artykuły:
- David A. Milner, Melvyn A. Goodale, Mózg wzrokowy w działaniu. Tłumaczenie: Grzegorz Króliczak Wyd. Naukowe PWN, Seria: Biblioteka Psychologii Współczesnej, 2008
- Crick Francis, Zdumiewająca hipoteza. Prószyński i S-ka, Warszawa 1997
- N. de Gelder, Widzący niewidzący, Świat Nauki, 6/2010 (popularny artykuł o ślepowidzeniu).
- Farah, M. J. (2004). Visual Agnosia, second edition. MIT Press.
- Fischer, J.T, Whitney, D. (2014). Serial dependence in perception. Nature Neuroscience. In Press.
- Hoffman, D. (2019). The Case Against Reality: Why Evolution Hid the Truth from Our Eyes. W. W. Norton & Company.
- Murzyn, E. (2008). Do we only dream in colour? A comparison of reported dream colour in younger and older adults with different experiences of black and white media. Consciousness and Cognition.
- Schwitzgebel E, Why Did We Think We Dreamed in Black and White? Studies in History and Philosophy of Science, 33, 649-660, 2002
- Voos, A., Cordeaux, C., Tirrell, J., & Pelphrey, K. (2013). Chapter 19 - Developmental Neuroscience of Social Perception. In J. L. R. Rubenstein & P. Rakic (Eds.), Neural Circuit Development and Function in the Brain (pp. 351–365). Oxford: Academic Press.
- Watanabe, M ... N Logothetis, Attention But Not Awareness Modulates the BOLD Signal in the Human V1 During Binocular Suppression. Science 334, 829-831, 2011
Cytowanie:
Włodzisław Duch, Wstęp do Kognitywistyki. Rozdz. B17: Percepcja wzrokowa. UMK Toruń 2024.