Percepcja: | Czucie | Słuch i Percepcja muzyki | Zmysł równowagi | Smak | Węch | Synestezje i ideastezje
Sygnały wysyłane przez receptory trafiają do obszarów pierwszorzędowych kory. Procesy samoorganizacji prowadzą do powstania w mózgu map pozwalających na rozróżnianie impulsów dochodzących od różnych receptorów. Omawialiśmy już ogólną organizację i podstawowe mechanizmy przetwarzania informacji, pora na bardziej szczegółowy opis w jaki sposób sens tej informacji jest analizowany i używany by podjąć odpowiednie działania.
Wtórna i trzeciorzędowa kora zmysłowa korzysta nie tylko z informacji otrzymanych z pierwszorzędowych obszarów, ale by nadać im znaczenie odwołuje się do pamięci, zapamiętanych bodźców i reakcji pozwalających na ich eksplorację.
Analizowaliśmy już przepływ informacji wzrokowej, teraz przyjrzymy się percepcji innych sygnałów zmysłowych.
Co dzieje się z informacją przez nie przesyłaną? Wiemy już, że percepcja służy działaniu, wiec najpierw z sygnałów zmysłowych trzeba wydobyć informacje przydatne do działania a potem przesłać je do płatów czołowych.
B16.1. Czucie i propriocepcja
Układ czuciowy jest unikalny, to jedyny układ, który nie jest zlokalizowany w niewielkim obszarze, tylko ma receptory rozrzucony po całym ciele: skóra jest naszym największym organem, każdy włosek połączony jest z receptorem, mamy też receptory w głębokich strukturach ścięgien, stawów i mięśni. Bez układu czuciowego niemożliwe by były precyzyjne ruchy. Jeśli mocno zmarznie nam twarz lub ręka, lub uszkodzony zostanie jakiś nerw, tracimy zdolność do kontroli mięśni w tym obszarze. Zatrucie nerwu u dentysty znacznie zimniejsza możliwości mimiki twarzy. Liczba włókien nerwowych przenoszących informację od receptorów na ciele człowieka oceniana jest na około 200-270 000 (Corniani i Saal 2020).
Część sygnałów z receptorów czuciowych wykorzystywana jest do automatycznych odruchów. Blokowanie przepływu tych sygnałów można wywołać lokalnie, uciskając lub masując bolące miejsce. Część sygnałów trafia do układu limbicznego, wpływając na aktywację wielu znajdujących się tam obszarów. Hamowanie dopływających stamtąd sygnałów możliwe jest dzięki pętlom wzgórzowo-korowym, mechanizmom uwagi, które synchronizując przepływ informacji pomiędzy różnymi obszarami pozwalają skupić się na wykonywanym zadaniu i ignorować sygnały czuciowe. Część sygnałów trafia do kory somatosensorycznej, czyli kory SI. Impulsy docierają tam od różnych receptorów, których aksony mają odmienne właściwości, a następnie różnych obszarów kory mózgu i układu limbicznego.
Aksony tworzą wiązki otoczone izolująca
wspólną otoczką (mieliną),
zwane nerwami.
Lista różnych nerwów jest długa.
Włókna nerwowe różnią się szybkością przekazywania sygnałów, zależnie od stopnia ich mielinizacji.
Nerwy dzieli się na 3 typy i wiele podtypów, klasyfikowanych w różny sposób - tu wymieniam tylko najważniejsze.
Włókna nerwowe typu Aα są dobrze zmienilizowane i przenoszą informację z prędkością do 120 m/sek.
Służą zarówno do przekazywania informacji zmysłowej jak i kontroli ruchu mięśni. Mogą przekazać sygnały bólowe z dowolnego miejsca ciała w ciągu 10 ms. Włókna przenoszące informację o czuciu i bólu są w rdzeniu kręgowym od siebie oddzielone aż do pnia mózgu. Nieco wolniej działają włókna receptorów zimna, ciepła, dotyku i sygnałów propriocepcyjnych.
Włókna typu B są słabiej zmienilizowane, przewodzą impulsy z szybkością do 14 m/sek, głównie z autonomicznego systemu nerwowego, gruczołów wewnętrznych.
Nerwy typu C pozbawione są mieliny, przekazują sygnały powoli (poniżej 2 m/sek). Takie włókna nerwowe przekazują informację o dotyku, swędzeniu, pieczeniu, temperaturze.
Nerwy czuciowe dochodzą przez projekcje z wzgórza do kory SI, w której mieszczą się obszary Brodmanna BA3, BA1 i BA2 (licząc od tyłu do przodu głowy). Najważniejszy jest obszar BA3, bo do niego dochodzą projekcje z wzgórza, neurony tego obszaru nie reagują na bodźce o innych modalnościach, lezje powodują brak odczuwania bodźców somatycznych, a pobudzenia prądem wywołują takie odczucia. Część obszaru B3 specjalizuje się w propriocepcji. Do obszaru B2 dociera informacja o kształtach i rozmiarze, a do obszaru BA1 o strukturze powierzchni (teksturze). Te pobudzenia mówią nam jak gładki jest jedwab lub jak chropowaty jest papier ścierny.
Zniszczenie kory SI powoduje zanik wrażeń czuciowych, jednak wrażenia bólu i temperatury po pewnym czasie mogą się pojawić, gdyż część projekcji ze wzgórza trafia do ciała migdałowatego i kory wyspy, co do pewnego stopnia umożliwia kompensację braku aktywacji kory SI.
Ilustracja wideo: szlaki czuciowe, czyli radiacja somatosensoryczna (tylko lokalnie).
|
Rozdział "Bezcielesna kobieta" w książce Oliver Sacksa "Mężczyzna, który pomylił swoją żonę z kapeluszem" (1996) opisuje rzadki przypadek kobiety, która utraciła propriocepcję w wyniku zapalenia nerwów czuciowych. Nie wiedziała, gdzie są jej nogi czy ręce, nazywając priopriocepcję "oczami ciała". Przypadek 19-letniego mężczyzny opisany przez Jonathana Cole w książce
"Losing touch" z powodu wirusowej infekcji po wielu latach ćwiczeń do pewnego stopnia odzyskał kontrolę nad ciałem. Obserwując i wyobrażając sobie swoje ruchy mu na stworzył sobie wewnętrzną mapę ciała pozwalającą na precyzyjną wizualizację tak by częściowo przywrócić normalne funkcje kontroli ciała. Zostało to pokazane w filmie
„Człowiek, który stracił ciało” (BBC Horizon 1988).
Informacja z grzbietowego szlaku wzrokowego dociera do pierwotnej kory czuciowej, ale aktywacje szybko znikają jeśli przestaje się patrzeć na położenie ciała.
To bardzo rzadkie przypadki, ale znacznie częściej zdarza się, że ludzie tracą czucie w samych palcach. Wówczas trudno im wykonywać takie czynności jak wiązanie sznurówek jeśli nie patrzą na swoje ręce. To przykład
astereognozji, czyli jednej z form agnozji czuciowej.
Czasami ludzie nie zauważają, że nie rozpoznają za pomocą dotyku przedmiotów lub nie potrafią wykonać pewnych czynności bo bezwiednie patrzą na swoje ręce. Podobnie jak skotoma w przypadku wzroku pokazuje to jak silnie sprzężona jest percepcja z kontrolą ruchów.
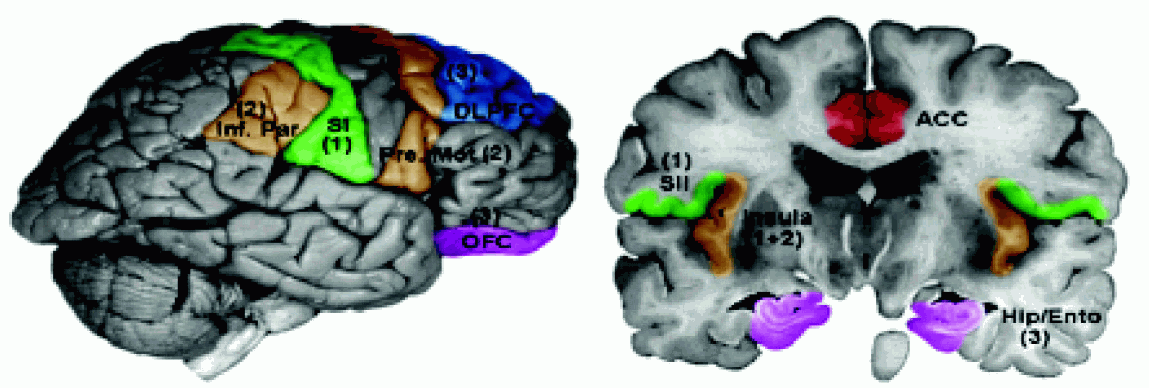
Diagram ilustrujący połączenia szlaków czuciowych pokazuje połączenia kory SI z drugorzędową korą czuciową SII, która łączy się z korą wyspy, ciałem migdałowatym, hipokampem, oraz pobliskimi obszarami ciemieniowymi BA5 i BA7, a przez brzuszny obszar ciemieniowy (VPC) z korą przedruchową.
Wtórna kora czuciowa SII leży w głębi bruzdy środkowej i bruzdy bocznej (BA 43), w pobliżu kory wyspy, chociaż niektórzy zaliczają do SII również płacik ciemieniowy dolny (obszar BA5), nieco z tyłu w stosunku do SI.
Obszar ten wyróżniono tylko u ludzi i małp naczelnych, jest to obszar kojarzeniowy dla wrażeń dotyku i bólu.
Komórki pobudzane są przez sygnały z większych obszarów ciała, z obu stron, bezpośrednio ze wzgórza, obszarów kory SI oraz spoidła wielkiego.
Integracja informacji z obu stron ciała oraz hipokampu pozwala na uczenie się rozpoznawania przez dotyk i transfer tej umiejętności z jednej ręki do drugiej.
Wtórna kora czuciowa SII zaangażowana jest również w interpretację emocjonalną sygnałów czuciowych.
Dalsza analiza bodźców czuciowych
Obszary BA5 i BA7 kory ciemieniowej, przylegające do kory somatosensorycznej traktowane są jako drugorzędowe obszary czuciowe, chociaż ich rola jest odmienna niż obszaru SII. Aktywacje kory SI pobudzają obszary ciemieniowe. Percepcja służy działaniu.
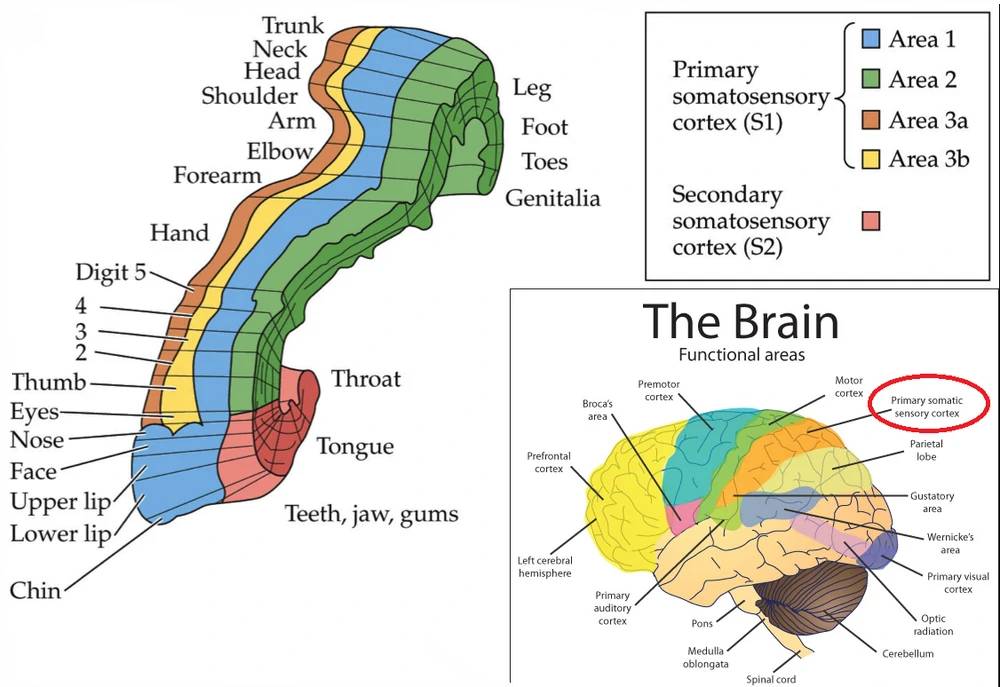
Odkrycie pod koniec lat 1930 przez Wildera Penfielda topograficznej organizacji kory SI przez długi czas wywołało przekonanie, że kora SI reprezentuje ciało w jednolity sposób. Dopiero pod koniec lat 1970 badania Jona Kaasa i jego współpracowników pokazały, że można tam wyróżnić cztery reprezentacje ciała, związane z różnymi receptorami.
Obszar BA1 otrzymuje sygnały ze skóry od receptorów, które się szybko adaptują, a obszar BA3b od adaptujących się powoli.
Obszar BA3a reaguje na receptory w mięśniach, a obszar BA2 na sygnały ze stawów i głębokich tkanek. Później odkryto jeszcze komórki czuciowe w korze ruchowej (BA4), tworzące piątą reprezentację. Czucie i ruch musi być silnie sprzężone by korygować ruchy w odpowiedni sposób.
Percepcja rozdziela wiec sygnały na różne typy pobudzeń, potrafimy rozróżnić dotyk palców u nogi od napięcia mięśni stopy, sygnały z ciała związane z wykonywanymi przez nas ruchami. Rozpoznajemy też siłę nacisku, kierunek ruchu, chropowatość powierzchni, temperaturę i ciężar unoszonych przedmiotów.
Te informacje tworzą całościowe wrażenie pozwalające na integrację na poziomie potrzebnym do podjęcia działania całego organizmu. Informacja staje się coraz mniej szczegółowa, pola recepcyjne coraz większe, sygnały z podobszarów kory SI zbierane są przez wielomodalne neurony w obszarze BA2, reagujące na wiele aspektów, które odczuwamy trzymając jakiś przedmiot w rękach.
Homunkulus ruchowy został szczegółowo przeanalizowany przy pomocy fMRI.
Okazało się, że
mapa ruchowa jest bardziej skomplikowana.
W korze M1 mamy obszary, które tworzą czuciowo-kognitywną sieć działania (somato-cognitive action network, SCAN), związaną z planowaniem złożonych ruchów. Większość kory wpływa na precyzyjne, izolowane ruchy naszych kończyn i ust, a SCAN integruje informacje dotyczące ruchu całego ciała, planów i celów działania z informacjami z autonomicznego układu nerwowego. Tą sieć można określić jako łączącą stany umysłu (plany, cele) i ciała (interocepcja i ruch).
Oprócz kilku
fragmentów kory M1 pomiędzy reprezentacją bioder/ramion/oczu,
sieć SCAN obejmuje wybrane regiony dodatkowej kory ruchowej (SMA), fragmenty wzgórza (VIM, CM), tylną część skorupy (putamen) i fragmenty móżdżku kontrolujące postawę. Ma też połączenia z brzuszną częścią kory ACC
(dACC, dorsal anterior cingulate cortex), które współpracują z obszarmi kory ciemieniowej w realizacji intencjonalnych ruchów, oraz fragmentami kory wyspy przetwarzającej sygnały czuciowe, bólu i interoceptywne sygnały z trzewi.
To mechanizmy rozwinięte u ludzi, pozwalające na integrację ruchów oczu i kończyn, używanie narzędzi, oddychanie i mówienie, utrzymywanie procesów aleostazy (czyli dynamicznej regulacji homeostazy). Sieć SCAN jest podstawą integracji, antycypacji zmian oddechu, aktywacji serca, przyspieszenia tętna, postury ciała i wstępnego pobudzenia mięśni, czy poczucia "motyli w brzuchu" wynikające z reakcji neuronów
enterycznego układu nerwowego
na adrenalinę. Planowanie, działanie i kontrola ciała są więc połączone we wspólnej rozległej sieci.
|
Głaskanie i łaskotki.
Głaskanie wywołuje przyjemne uczucia u wielu zwierząt i ludzi. Czym wyróżnia się
łaskotanie
i czemu nie możemy się sami połaskotać?
Badania S. Blakemore przy użyciu głaskającego robota pokazały, że regularne głaskanie nie wywołuje wrażenia łaskotania. W tej sytuacji wiemy, czego się możemy spodziewać. Jednakże nieprzewidywalne zachowanie głaskającego robota wywołuje wrażenie łaskotania. Można samemu wykonać takie ćwiczenie z drugą osobą, głaskająca rękę za zasłoną w sposób przewidywalny lub przypadkowy - jakie wówczas będą Wasze wrażenia?
Badania łaskotania dały
zaskakujące wyniki: ludzie budzenie w fazie REM byli w stanie siebie łaskotać, być może zanikła chwilowo zdolność do przewidywania swojego działania. W snach
świadomie kontrolowanych
(lucid dreams) nie udaje się nikomu być łaskotanym. Interpretacja tych wyników nie jest jasna.
Czy nie udaje się w nich utworzyć postaci na tyle niezależnych by działały w sposób nieprzewidywalny? Czy poczucie sprawczości w świadomym śnie, który wymaga przygotowania, jest zbyt silne? Część brzuszna jest schowana w głębi bruzdy, a część grzbietowa bardziej na zewnątrz.
Stymulacja tego obszaru wywołuje nie tylko wrażenia czuciowe, ale też pobudza korę ruchową. Stymulacja części grzbietowej wywołuje ruchy całego ciała, przypominające podskoki i wspinanie, a części brzusznej wskazywanie na okolice bliskie ciała lub w kierunku ust. Informacja, która powstaje w korze SII jest już wystarczająca by podjąć szybkie działanie. Informacja w obszarach BA5 i 7 kory ciemieniowej wystarcza by zidentyfikować miejsce i części ciała, które mogą posłużyć by do niego sięgnąć. Kora SII wykorzystuje topograficzną organizację kory ruchowej by uruchomić odpowiednie części ciała. Integracja informacji wzrokowej i czuciowej to podstawa działań senso-motorycznych, czyli postrzeżenia i działania. Brakuje tylko oceny poznawczej pozwalającej na bardziej złożone plany działania.
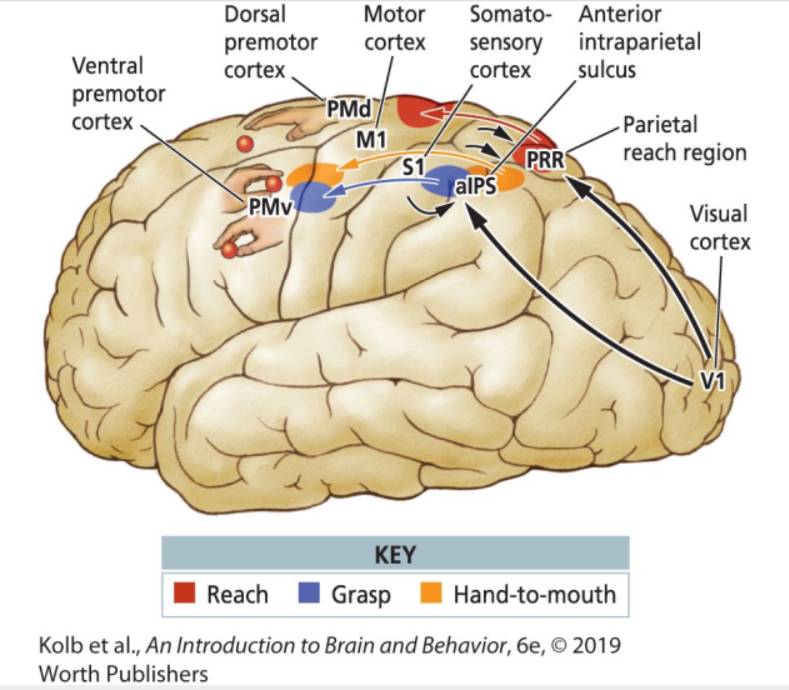
|
Mamy trzy rodzaje ruchów rąk, wymagających odmiennej interpretacji bodźców czuciowych i innego sterowania. Sięganie i łapanie związane jest z pobudzeniami części grzbietowej i brzusznej, które otrzymują sygnały z kory SI oraz kory wzrokowej. Ruchy związane z wkładaniem jedzenia do ust aktywowane są przez obszar pośredni SII, który nie otrzymuje informacji z kory wzrokowej. Tylko te ruchy nie wymagają koordynacji wzrokowej, drugorzędowa kora somatosensoryczna nauczyła się metodą prób i błędów jak pobudzić korę ruchową by trafić do ust. Jak widać nasze ruchy składają się z elementarnych działań, aktywacji specyficznych programów ruchowych w wyniku percepcji. Kora ciemieniowa, somatosensoryczna i ruchowa ściśle ze sobą współpracuje.
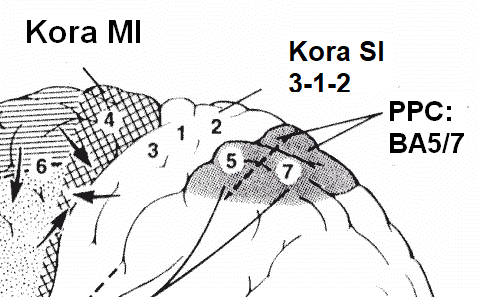
|
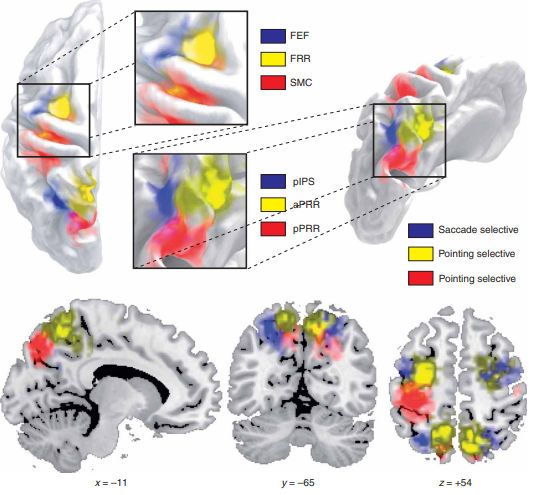
Kora ciemieniowa integruje informacje wzrokowe na temat obserwowanego ruchu, a ta informacja służy do sterowania mięśniami kontrolującymi ruch sakadyczny gałek ocznych i kończyn. W obszarze przyśrodkowej kory ciemieniowej zwanej
przedklinkiem zidentyfikowano regiony
(Tosoni i inn. 2008) oznaczone skrótem
aPRR (anterior parietal reach region, przedni ciemieniowy region chwytania, kolor żółty), oraz
pPRR (posterior parietal reach region, tylny ciemieniowy region chwytania, kolor czerwony), oraz region związany z sakadami, oznaczone skrótem
pIPS (posterior intraparietal sulcus,
bruzda śródciemieniowa tylna, kolor niebieski).
Uszkodzenia obszarów leżących w głębi
przyśrodkowej kory ciemieniowej (LIP), części kory środciemieniowej IPS, wywołuje niezdolność do chwytania przedmiotów, np. lecącej piłeczki. Obszar LIP zawiera topograficzną mapę obiektów istotnych z punktu widzenia bieżących możliwości działania, odwołania do specyficznych ruchów ciała i oczu, kategoryzację obiektów z punktu widzenia możliwych działań, spodziewane nagrody. LIP odpowiada więc za integrację informacji wzrokowo-przestrzennych, poznawczych i ruchowych, z mechanizmami uwagi.
W korze ruchowej zidentyfikowano trzy regiony (ROI)
selektywnie reagujące na sygnały z odpowiednich obszarów ciemieniowych. Pierwszym z nich jest obszar
FEF (frontal eye field, czołowe pole ruchowe gałek ocznych), powiązany z regionem pIPS (obydwa zaznaczone na niebiesko). FEF to jeden z kluczowych obszarów kontrolujących ruch naszych oczu, zarówno wolicjonalny, śledzenia jakiegoś obiektu (cele wyznacza kora przedczołowa), jak i związany z orientacją (sygnały z pIPS). Aktywacja tego regionu pojawia się już po 45 ms od wzrokowego bodźca i po 24 ms od bodźców akustycznych, zmieniając swoją charakterystykę z zależności od dźwięku już po 30-60 ms. FEF i pIPS wysyłają rezultaty swojej analizy do wzgórków czworaczych górnych. Są to struktury śródmózgowia, w których tworzy się mapa istotności bodźców wzrokowych, a w dolnych wzgórkach czworaczych słuchowych, docierają tam też bodźce z różnych zmysłów. Aktywność tego obszaru ma duży wkład w możliwości skupiania się,
unikania rozproszenia.
Dwa pozostałe ROI kory ruchowej, oznaczone jako
FRR (czołowy obszar chwytania) oraz
SMC (sensory motor cortex, kora czuciowo-ruchowa), czyli część kory ruchowej silnie sprzężonej z korą czuciową.
Eksperymenty pokazały, że aktywność w tych regionach nie jest związana z samym pojawieniem się bodźców wzrokowych, tylko z połączeniem sygnałów zmysłowych z uwarunkowanym przez kontekst działaniem. Zmysły integrują wskazania pozwalające na podjęcie określonych działań, to od tego zależy aktywacja określonych obszarów kory ruchowej.
Postrzeżenie-działanie jest silnie sprzężone nie tylko w przypadku prostych reakcji sensomotorycznych, ale też bardziej złożonych decyzji, które sobie uświadamiamy i uznajemy za wolicjonalne, chociaż samo ich wykonanie nie wymaga świadomości. To przykład zachowań, w których świadomość pełni rolę epifenomenu, tzn. występuje prawie jednocześnie ale nie wywiera wpływu na dane zdarzenie.
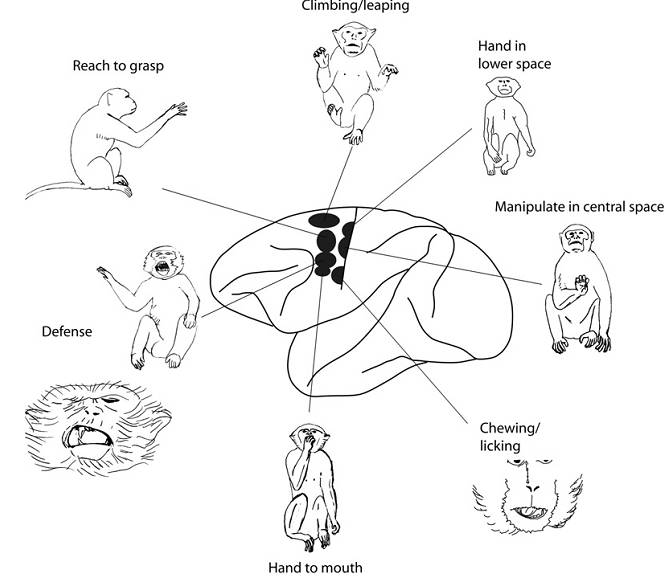
|
Dokładne badania reakcji kory ruchowej na różne rodzaje ruchów pozwoliły odróżnić nie tylko sięganie i chwytanie, ale kilka innych rodzajów ruchów. Wydaje się, że jest tu wspólny mechanizm integrujący percepcję wzrokową, czuciową i specyficzne ruchy kończyn.
|
Uszkodzenia kory somatosensorycznej
Uszkodzenia kory SI zaburzają nawet proste czynności, takie jak zdolność do identyfikacji przedmiotów za pomocą dotyku, określenie kierunku ruchu ręki lub palca, czy odczucie siły nacisku. Takie upośledzenie precyzyjnych celowych ruchów nazywa się
apraksją.
Istnieje wiele form apraksji, w zależności od rodzaju uszkodzeń. Apraksja ruchowa może się objawiać niezdolnością do wykonywania precyzyjnych ruchów, np. pisania, ale niektóre uszkodzenia prowadzą do apraksji wyobrażeniowej, w której pacjentom trudno jest wykonać polecenia związane z koniecznością zaplanowania ruchów.
Istnieje też apraksja związana z mową, ruchem oczu, zdolnością do rysowania, ruchów twarzy i wielu innych form.
Specyficzne zaburzenia pokazują jak wyspecjalizowane są obszary kory, kontrolujące percepcję i ruch.
Rola informacji analizowanych w drugorzędowej korze czuciowej górnej części kory ciemieniowej wskazuje na możliwość dysocjacji pomiędzy działaniem a możliwością rozpoznania i opisu tego co robimy. To właśnie stało się przyczyną rozwinięcia teorii dwóch strumienie informacji, działania i percepcji, przez Milnera i Goodale (1992).
Agnozja (gr. a=negatywne, gnosis=wiedza) to niezdolność lub zaburzenie rozpoznania lub identyfikacji informacji zmysłowej. Większość agnozji należy rozpatrywać na wyższym poziomie analizy, nie tylko receptorów lub pierwszorzędowej kory analizującej dane zmysłowe.
Agnozja czuciowa lub astereognozja to niezdolność do rozpoznania przedmiotów za pomocą dotyku. Jest to niezależne od zaburzeń czucia upośledzenie rozpoznawania kształtu przy prawidłowym rozpoznawaniu jego wielkości, ciężaru lub faktury powierzchni.
Optyczna ataksja to niezdolność do chwytania przedmiotów pomimo prawidłowej analizy wzrokowej, np. nie można wlać wody do szklanki. To jeden z objawów rzadkiego zespołu Bálinta, w którym mogą też wystąpić inne zaburzenia, takie jak
symultanagnozja,
czyli problem z rozpoznawaniem relacji pomiędzy wieloma obiektami, które stanowią całościową strukturę. Uwaga jest skupiona na małej przestrzeni i postrzegane są pojedyncze obiekty, więc przeniesienie wzroku z łyżki na zupę powoduje, że łyżka znika.
Symultanagnozja może być wynikiem uszkodzenia szlaku brzusznego bądź grzbietowego. W przypadku obustronnych uszkodzeń szlaku grzbietowego (kory potyliczno-ciemieniowej) postrzeganie zawężone jest do jednego pola, zanika świadomość istnienia czegokolwiek poza nim. W efekcie pacjenci mają trudności z poruszaniem się, ale też z czytaniem. Postrzeganie przedmiotów w ruchu jest również utrudnione. Uszkodzenia obszaru brzusznego w okolicach lewej strony dolnej części połączenia potyliczno-skroniowego (inferior occipito-temporal junction) nie zaburza tak silnie widzenia kolejnych przedmiotów, ale nie pozwala widzieć całej sceny, wymagając skupiania uwagi na jej kolejnych elementach. To powoduje również problemy z czytaniem
(nazywa się to "czystą aleksją", bo inne funkcje językowe są w normie, np. nazywanie przedmiotów, powtarzanie wyrazów, rozumienie słów czy pisanie). Pacjenci z brzuszną symultanagnozją poruszają się bez obijania o przedmioty w pomieszczeniu.
W obu przypadkach wielkość obserwowanych elementów nie ma znaczenia, ważna jest tylko ich liczba.
Są to rzadkie formy agnozji, pokazują też, jak ważnej est posiadanie dwóch półkul mózgowych, dzięki czemu możliwa jest kompensacja uszkodzeń po jednej stronie mózgu.
SSA, dodatkowa kora czuciowa, znajduje się w płaciku ciemieniowym górnym (obszary 5 i 7 Brodmanna).
Obszary czuciowe w płacie ciemieniowym mają złożone funkcje:
- Stereognozja to identyfikacja dotykowa obiektów.
- Możliwe jest rozpoznawanie kształtów, rozmiarów i tekstur.
- Do utworzenia się zdolności sterognozji potrzebna jest nauka oparta na integracji wrażeń różnych modalności, szczególnie wzrokowo-czuciowych.
- Dermoleksja to zdolność do rozpoznawania kształtów (np. znaków) kreślonych na skórze np. palcem. Rozpoznawanie znaków pisma na korze nazywa się grafestezją.
Kora czuciowa, podobnie jak kora ruchowa, zdolna jest do znacznej reorganizacji. Powstały szczegółowe modele określające w której warstwie tworzy się szczegółowa reprezentacja obszarów ciała, oraz jak zmienia się ta reprezentacja na skutek stymulacji (np. częstego używania kciuka do pisania wiadomości na telefonie), czy tworzenia się reprezentacji w przypadku kończyn fantomowych.
Kora czuciowa ma trochę neuronów ruchowych (tylko dwie synapsy dzielą ją od kontroli mięśni), a kora ruchowa neuronów czuciowych.
Czucie zewnętrzne to analiza sygnałów z narządów zmysłów.
Czucie głębokie to analiza sygnałów proprioceptywnych z mięśni, stawów, ścięgien, błędnika, informujące o położeniu i stanie ciała.
Czucie wewnętrzne, interoceptywne, analizuje informację z gruczołów i trzewi.
Połączenie kory SI i SSA umożliwiają przepływ informacji pomiędzy lokalizacją wzrokową a pobudzeniem czuciowym.
Najlepszym przykładem tego współdziałania są kończyny fantomowe.
Kończyny fantomowe są wynikiem silnego odczucia obecności amputowanych kończyn, pojawiające się u 60-80% osób po amputacji.
Często jest to odczucie bolesne, czasami mogą sie pojawić wszelkie inne wrażenia czuciowe: ciepło/zimno, swędzenie, ucisk, łaskotanie, wrażenie ruchu i próba chwytania nieistniejącą kończyną.
Odpowiedzialna jest za to reorganizacja kory czuciowej wynikająca z pobudzeń z okolic ciała, które mają zbliżone pola projekcyjne w obszarze SI, słabe połączenia normalnie maskowane przez odczucia płynące z brakującej części ciała.
Takie odczucia nie pojawiają się w przypadku porażenia kończyn dolnych (paraplegii).
Halucynacje czuciowe
Kończyny fantomowe są specyficznym rodzajem halucynacji związanych z pobudzeniem kory czuciowej.
Nasze wrażenia to interpretacja stanów różnych obszarów mózgu, pobudzanych przez dane zmysłowe lub wynikające z wewnętrznych pobudzeń. Dlatego mogą się oczywiście zdarzać halucynacje różnych rodzajów.
Cenestezja to odczucia i wrażenia odbierane z wnętrza ciała.
Cenestezyjne halucynacje oralne to wrażenia gazu wydobywającego się z ust, drutów albo ślimaka, który jest na dziąsłach.
Kończyny niechciane (dysmorfia ciała) to
silny wstręt do własnego ciała, poczucie obcości, dość częste, 1:50 osób. Skrajne przypadki zaczynają się w dzieciństwie, "życie w ciele, które nie jest własne".
To zaburzenie traktowane jest jako obsesja, pacjenci walczą o prawo
do amputacji kończyn (apotemnofilia).
Kilku osobom w Wielkiej Brytanii obcięto kończyny (pokazuje to film BBC Horizon
'Complete Obsession').
To zjawisko należy do szerszej kategorii określanej jako xenomelia lub
dysforia integralności ciała.
Mentalne wyobrażenie nie zawsze pasuje do fizycznej budowy ciała, już w wczesnym dzieciństwie pojawia się silne pragnienie pozbycia się części ciała.
Co może być przyczyną nieprzyjemnych odczuć czuciowych?
Sprzężenie obszarów interpretujących informację z kończyn w obszarze SII z ciałem migdałowatym może wywołać nieprzyjemne odczucia, możliwe są też inne odczucia, np. podniecenie seksualne. W jednym z badań na 16 kandydatach do amputacji nogi stwierdzono, że górny płacik ciemieniowy (SPL), część kory ciemieniowej (PPC) pełniąca rolę drugorzędowej kory somatosensorycznej, jest cieńsza i zawiera mniej szarej materii. Wiadomo, że lezje w tym obszarze wywołują uczucie zaniku części ciała. Jeśli nie ma poczucia, że mogę po coś sięgnąć czy coś chwycić to widać nie mam czym. Również brzuszna część kory przedruchowej (vPMC) uczestniczy w budowaniu obszaru ciała. Te obszary mają też
mniej połączeń funkcjonalnych.
Nie ma dobrych metod leczenia, można próbować pobudzania nerwów czuciowych w połączeniu z przyjemnymi bodźcami.
Gdzie jestem? Skąd to wiem? Nauczyłem się korelować wrażenia wzrokowe i czuciowe, wzrok dostarcza nam większość informacji do kory ciemieniowej, tworząc egocentryczną i allocentryczną mapę przestrzeni.
Wyjaśnia to film Henrika Ehrssona z TEDx:
What if we could leave our body and have a new one?
Impulsy dostarczane za pomocą mikroelektrod umieszczonych w obszarach somatosensorycznych SI (zwykle można to badać na zwierzętach lub sparaliżowanych osobach) wystarczają by stworzyć odczucia dotyku i wykorzystać je jako sygnały pomagające manipulować zewnętrznymi urządzeniami, np. wirtualnymi kończynami. Małpy po paru tygodniach treningu osiągnęły poziom rozróżniania bodźców czuciowych z sztucznej ręki porównywalny z prawdziwym. Badania na szczurach, którym w obszar czuciowy wibrysów wszczepiono elektrody przesyłające sygnały z detektorów podczerwieni pokazały, jak ta informacja pozwoliła im zintegrować widzenie w podczerwieni jako zupełnie nowy zmysł (Lebedev i Nicolelis, 2017).
Taka substytucja zmysłów stwarza to możliwości całkowitej symulacji wrażeń w wirtualnej rzeczywistości. Więcej na ten temat w rozdziale na temat ruchu i substytucji zmysłów.
Percepcja bólu
Odczuwanie bólu jest niezwykle ważnym zagadnieniem. To najczęstsza przyczyna wizyt u lekarzy. Ból jest sygnałem, że dzieje się coś złego, im groźniejsza sytuacja tym większy ból. Ludzie, którzy nie odczuwają bólu zwykle nie żyją długo ... bez bólu nie wiem, czy ułożenie ciała jest właściwe, czy nie jest ono zbytnio obciążone lub skręcone i w efekcie doprowadzimy do jego deformacji.
Około 8 różnych typów włókien nerwowych dostarcza do mózgu informacje na temat bólu, pobudza wydzielanie się hormonów, wywołujących stany zapalne, puchnięcie i zaczerwienienie uszkodzonych miejsc. Liczne ścieżki sygnalizujące ból uniemożliwiają chirurgiczne zabiegi i utrudniają zwalczanie bólu.
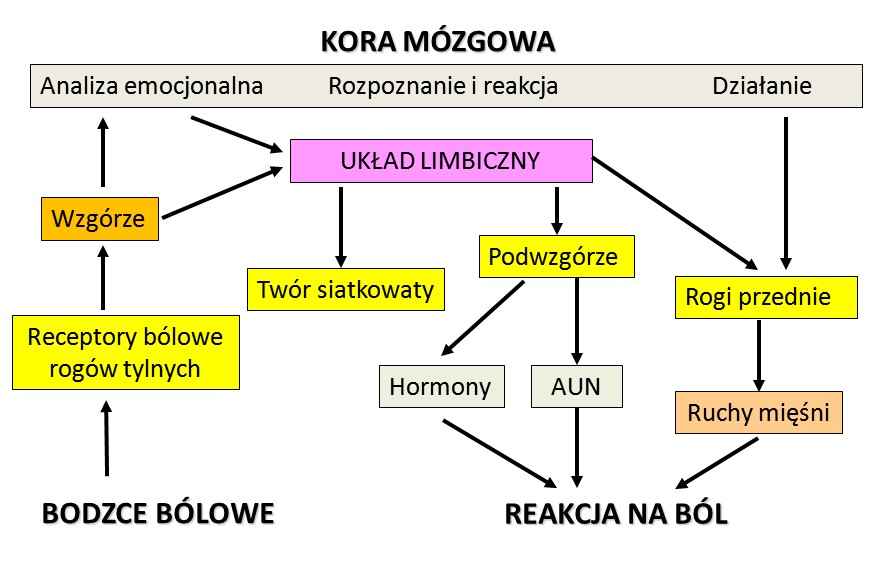
|
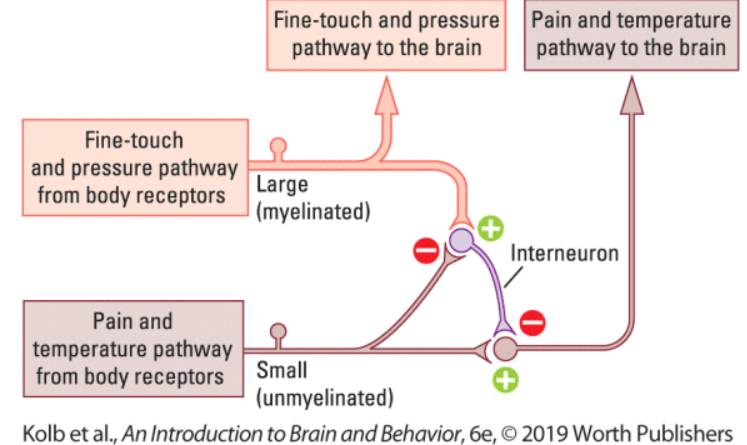
|
Teoria "bramkowania bólu" ( Melzack i Wall, 1965) zakłada, że pomiędzy włóknami przewodzącymi sygnały bólowe i czuciowe na poziomie rdzenia kręgowego są interneurony hamujące wzajemne pobudzenia. Ból chwilowo znosi czucie ale też bodźce czuciowe, takie jak pocieranie, masaż, ucisk, akupunktura, podgrzewanie czy inne zabiegi ograniczają przesyłanie impulsów bólowych. Lokalnymi neurotransmiterami oddziałującymi na interneurony są opiody.
W przypadku zabiegów chirurgicznych, czasami też w czasie porodu, stosuje się
znieczulenie zewnątrzoponowe,
wstrzykiwanie morfiny do przestrzeni zewnątrzoponowej kanału kręgowego. To pobudza interneurony, które hamują doznania bólowe.
Jest to jednak znieczulenie a nie terapia przewlekłego bólu.
Efekty mrowienia i drętwienia, zwane fachowo
parastezją, mogą mieć bardzo liczne przyczyny, ale wspólny mechanizm: dłuższy ucisk ogranicza przepływ informacji przez grubsze, zmienilizowane aksony, które dostarczają informacji o dotyku i nacisku, ale nie blokują cieńszych włókien typu C niosących informacje o temperaturze i bólu. Zwykle ćwiczenia fizyczne zwiększają ukrwienie przywracając normalne czucie.
Mechanizm hamowania może dotyczyć interneuronów nie tylko w rdzeniu kręgowym. Neurony
istoty szarej okołowodociągowej (PAG), otaczającej kanał wypełniony płynem mózgowo-rdzeniowym w śródmózgowiu, mogą hamować sygnały bólowe wpływając na jądra w pniu mózgu, modulując ilość produkowanych tam neurotransmiterów, hamując w ten sposób przepływ informacji do wzgórza. Głęboka stymulacja mózgu za pomocą elektrod implantowanych w PAG jest jedną z metod terapii lekoodpornego bólu.
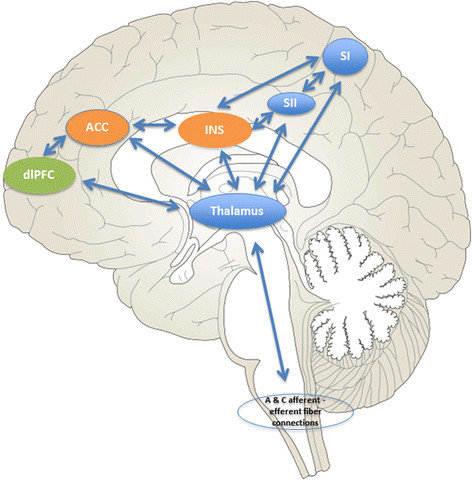
|
Bodźce bólowe nie muszą przechodzić przez dokładniejszą interpretację na poziomie kory by wywołać mobilizację i reakcję organizmu. "Macierz bólu" to zbiór obszarów, które wymieniają między sobą informację w przypadku sygnałów bólowych. Każde wrażenie, jakie odczuwamy na poziomie mentalnym, jest wynikiem procesów zachodzących w wielu obszarach mózgu, przepływu informacji, który nasze mózgi nauczyły się interpretować i wyrażać za pomocą reakcji behawioralnych i komentarzy słownych.
Niebieskim kolorem zaznaczono obszary odpowiedzialne za rozpoznanie miejsca i rodzaju sygnałów czuciowych.
Kolorem pomarańczowym (ACC, INS) te obszary, które związane są z emocjami i motywacją do działania.
Przednia część kory zakrętu obręczy (ACC) współpracuje z ciałem migdałowatym i innymi strukturami układu limbicznego.
Grzbietowo-boczna część kory przedczołowej (dlPFC), zaznaczona na zielono, pozwala na ocenę i interpretację bodźców i planowanie działania.
Organy wewnętrzne - tętnice i żyły, serce i nerki, mają receptory bólu ale nie mają swoich wydzielonych ścieżek sygnałowych. Z ewolucyjnego punktu widzenia precyzyjna informacja o bólu wewnętrznych organów była nieprzydatna, nie było możliwości odpowiedniej interwencji. Sygnały bólowe wynikające z interocepcji mieszają się w rdzeniu kręgowym z sygnałami czucia somatycznego. W ten sposób powstaje ból rzutowany, odczucia bólu na powierzchni ciała wywołane przez sygnały z wewnętrznych organów. Ból serca może spowodować drętwienie lewego ramienia, ból nerek przenosi się na plecy, a receptory bólowe w żyłach i tętnicach mózgu powodują ból głowy.
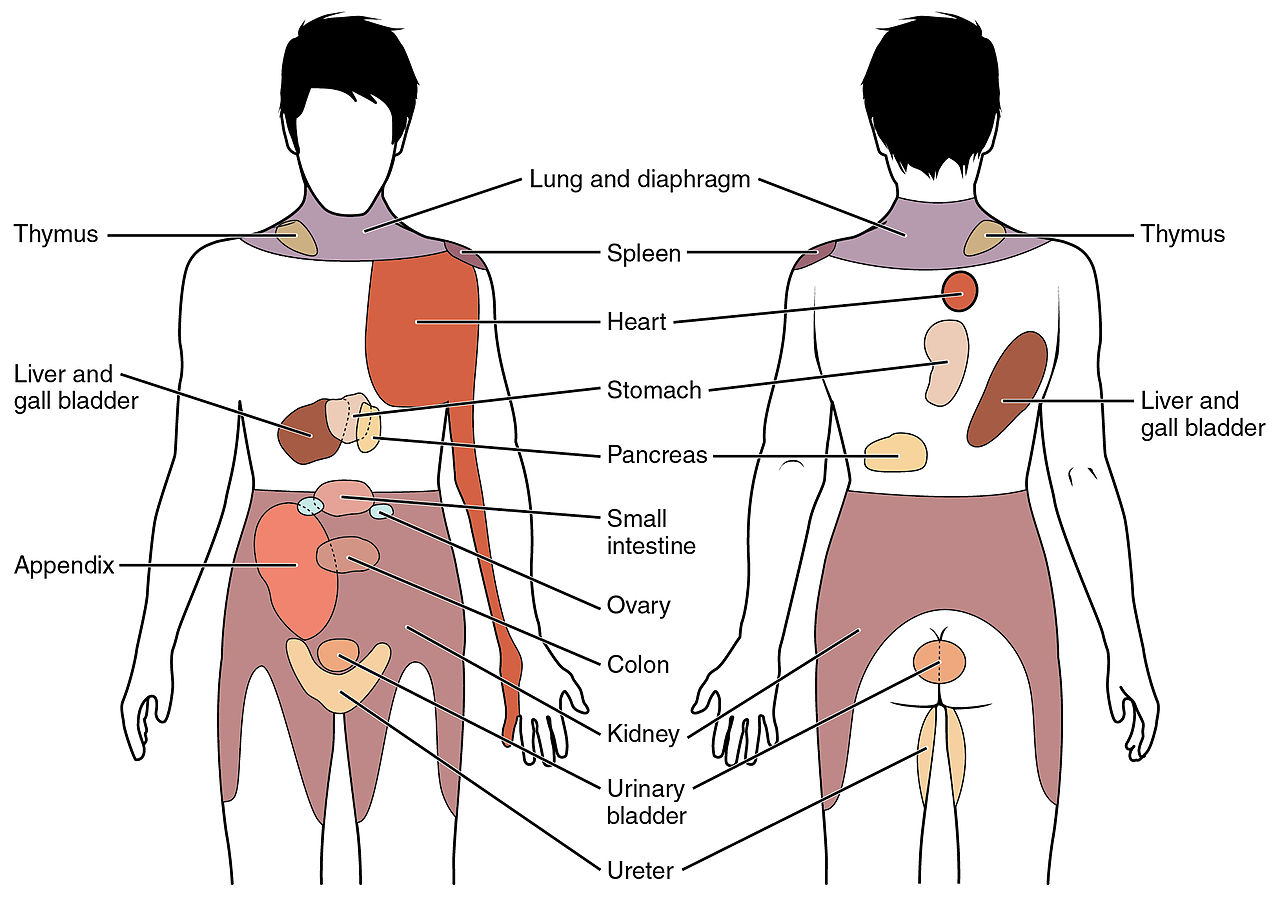
|
Swędzenie zwraca uwagę na potrzebę oczyszczenia skóry.
Można je zredukować
pobudzając receptory bólowe przez tarcie, nacisk, ciepło czy drażnienie prądem.
Drapanie blokuje przesyłanie sygnałów o swędzeniu przez rdzeń kręgowy. Drapanie może usunąć pasożyty lub czynniki chemiczne wywołujące swędzenie a powstałe zadrapania
powodują stan zapalny
zwalczający szkodliwe czynniki na skórze.
Swędzenie może pojawić się bez zewnętrznej przyczyny, z powodu pobudzenia nerwów.
Układy sensoryczne działają na granicy pobudzenia - zwiększa to ich czułość (podobnie jak w przypadku rezonansu stochastycznego grat tu rolę szum neuronalny) ale czasami powoduje fałszywe alarmy. np. swędzenie lub nadwrażliwość skóry.
Uszkodzenia drugorzędowej kory czuciowej jak i pobliskiej kory wyspy powodują
asymbolię bólu,
brak cierpienia (reakcji psychicznych) na ból, lub wywołują
stały ból piekący (kauzalgia).
Możliwa jest dysocjacja pomiędzy
wrażeniem bólu a reakcją emocjonalną, pomimo świadomości intensywności i umiejscowienia bodźca, który normalnie wywołuje ból.
Analgezja wrodzona to niezdolność do odczuwania bólu, pomimo odczuwania dotyku, ciepła, zimna i łaskotania.
Nie zachodzi przekazywanie impulsów bólowych do mózgu z powodu mutacji genu SCN9A, którego białka tworzą
sodowy kanał jonowy Nav1.7 w
receptorach bólowych (nocyceptorach)
jak i receptorach węchowych, stąd takie osoby cierpią również na
wrodzoną anosmię.
Łatwo się wówczas trwale uszkodzić nawet nie zauważając ...
Są różne
rodzaje analgezji, brak postrzegania bodźców bólowych - całkowita niewrażliwość, oraz obojętność, czyli brak reakcji pomimo odczuwania bodźca (jak napisała jedna z osób, "uwielbiałem dźwięk swojej skwierczącej skóry").
Damasio podaje opisy przypadków osób po operacji w okolicach kory zakrętu obręczy lub kory wyspy, po których pacjent nadal odczuwał ból, ale zniknęło cierpienie.
Możliwa jest opóźniona reakcja na ból, reakcja następująca wiele minut po ukłuciu (tak jest w przypadku późnego stadium syfilisu).
Niektóre środki przeciwbólowe obniżają aktywność kory przedniego zakrętu obręczy.
Ból wieńcowy wiąże się z pobudzenie tego samego obszaru.
B16.2 Słuch: drogi słuchowe, zaburzenia słuchu

|
Wikibook ś
Sensory Systems/Auditory System omawia budowę ucha, oraz kodowanie wysokości dźwięku.
Podsumowanie własnościach
słuchu jest tutaj.
Nie będziemy się tu zajmować
psychoakustyką, ani różnymi zjawiskami fizycznymi, związanymi z działaniem aparatu słuchowego, czy też fizjologią słuchu.
Fala zmian ciśnienia wywołuje wibracje, które postrzegamy jako dźwięk. Amplituda tej fali decyduje o intensywności zmian ciśnienia, a moc (natężenie dźwięku) jest proporcjonalna do kwadratu amplitudy. Zgodnie z prawem
Webera-Fechnera głośność dźwięku i intensywności innych wrażeń (ciężar, jasność, zapach) postrzegamy w skali logarytmicznej. Postrzegamy różne bodźce w bardzo szerokim zakresie zmian, wyczuwamy małe zmiany, ale jeśli bodziec jest silny to mała zmiana jest trudna do zauważenia. Nie postrzegamy więc bezwzględnych wartości tylko zmiany porównywalne do wielkości bodźca.
Formalnie mierzymy takie zmiany w
decybelach (dB), czyli jednostkach, które są 1/10 Bela (od wynalazcy telefonu Alexandra Grahama Bella). Jeden Bel odpowiada mocy 10 razy większej od progowej, To oznacza, że jeśli natężenie dźwięku wzrośnie o 10 razy to ocenimy ten wzrost na 10 dB, a jeśli wzrośnie 100 razy (czyli amplituda zmian ciśnienia wzrośnie 10 razy) to jest to wzrost o 20 dB.
Próg czułości, czyli poziom 0 dB, to zmiana ciśnienia atmosferycznego o jedną miliardową część, a próg bólu to zmiana rzędu 10 bilionów (1013, 130 dB) razy większa! Szept to 20 dB, normalna rozmowa koło 50-60 dB, hałaśliwa ulica 70, a głośny koncert może osiągnąć ponad 110 dB, czyli blisko ryku silników samolotu, ocenianych na 120 dB. Smartfony mają aplikację pozwalającą mierzyć poziom hałasu.
Około 16-20 tysięcy włosków w uchu środkowym (komórek rzęsatych) reaguje na fale dźwiękową umożliwiając rozróżnienie około 1500 wysokości dźwięków. Najniższy dźwięk fortepianu to A2 = 27,50 Hz, a najwyższy c5 = ok. 4186 Hz (88 klawisz).
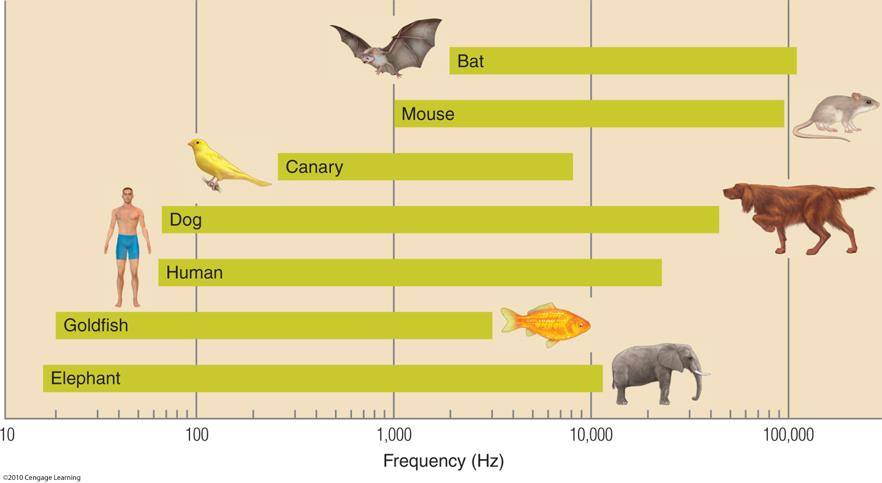
Przy dźwięku 1000 Hz możemy zauważyć różnice częstotliwości rzędu 1 Hz. Barwa dźwięku zależy od tego, jak skomplikowany jest jego przebieg, czyli na jakie częstotliwości możemy go rozłożyć. Różne zwierzęta słyszą dźwięki w różnym zakresie.
Postrzegana głośność zależy od częstotliwości dźwięku. Ewolucja dostroiła naszą wrażliwość do dźwięków mowy, największa czułośc słuchu wypada w okolicach 2-3 000 Hz.
|
Drogi słuchowe
Skupimy się nad ścieżkami transmisji informacji o zmianach ciśnienia. Omawialiśmy już receptory w uchu wewnętrznym odpowiedzialne za przetwarzanie niezwykle małych zmian ciśnienia na impulsy neuronowe. Co się z nimi dzieje dalej?
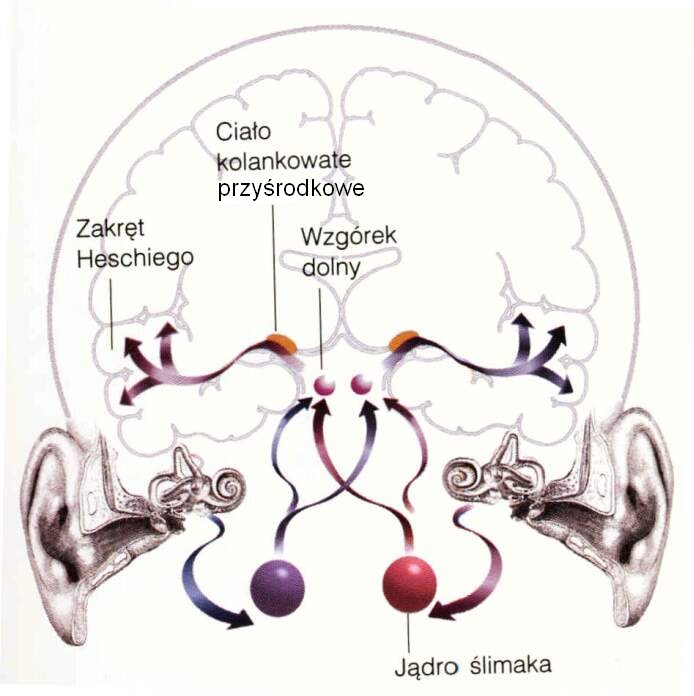
a stamtąd do wzgórza i pierwotnej kory słuchowej. |
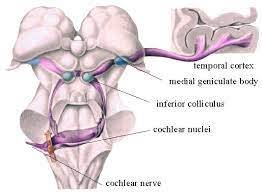
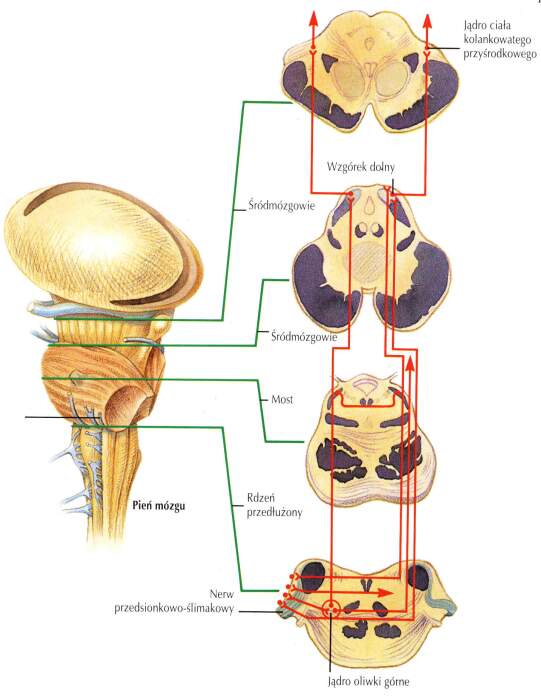
Sygnał z ślimaka wędruje najpierw do
jąder ślimakowych
(mają dwie części, brzuszną i grzbietową). Reakcje znajdujących się tam neuronów są bardzo złożone, nawet dla słabych sygnałów dostrojone do określonych częstotliwości. Pomaga to w lokalizacji źródła dźwięku w pionie.
Sygnały z każdego jądra ślimakowego trafiają następnie do jąder górnych oliwek po obu stronach, gdzie komórki specjalizują się w obliczaniu różnic czasu dotarcia dźwięku, dzięki porównaniu amplitudy i fazy sygnału, czyli różnicy intensywności i czasu narastania sygnału z każdego ucha.
Dla dźwięków o wysokiej częstotliwości, powyżej 1.6 kHz, najważniejsza jest różnica głośności (Interaural level differences, ILD), bo wówczas długość fali dźwiękowej jest mniejsza niż odległość między uszami, głowa tłumi nieco dźwięk dochodzący do dalszego ucha. Różnica czasu (interaural time differences, ITD) jest ważna dla niskich dźwięków, które zawijają się wokół głowy i odbierane są z podobną intensywnością.
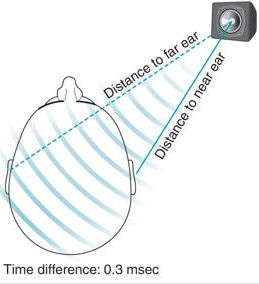
i lokalizacja źródeł. |
Dźwięk wędruje wokół głowy przez około 700 mikrosekund (μs), ale człowiek może zauważyć niesłychanie małe różnice nawet koło 10 μs. To zaledwie 1/10 cyklu dźwięku o częstotliwości 10 kHz, a więc bardzo wysokiego. Ten mechanizm pozwala na precyzyjną lokalizację źródeł dźwięku w poziomie, co pozwala na ustalenie kierunku wzroku z dokładnością prawie 1 stopnia, ale wymaga to treningu. Niektóre zwierzęta, np. sowy polujące nocą, są w stanie namierzyć gryzonie z dużej odległości. Wiele zwierząt potrafi też poruszać uszami, kierujące je w stronę źródeł dźwięku. Jądra ślimakowe i oliwkowe przetwarzają sygnały słuchowe pozwalające na orientację. Kształt ucha też pomaga w określeniu kierunku, skąd nadchodzi dźwięk.
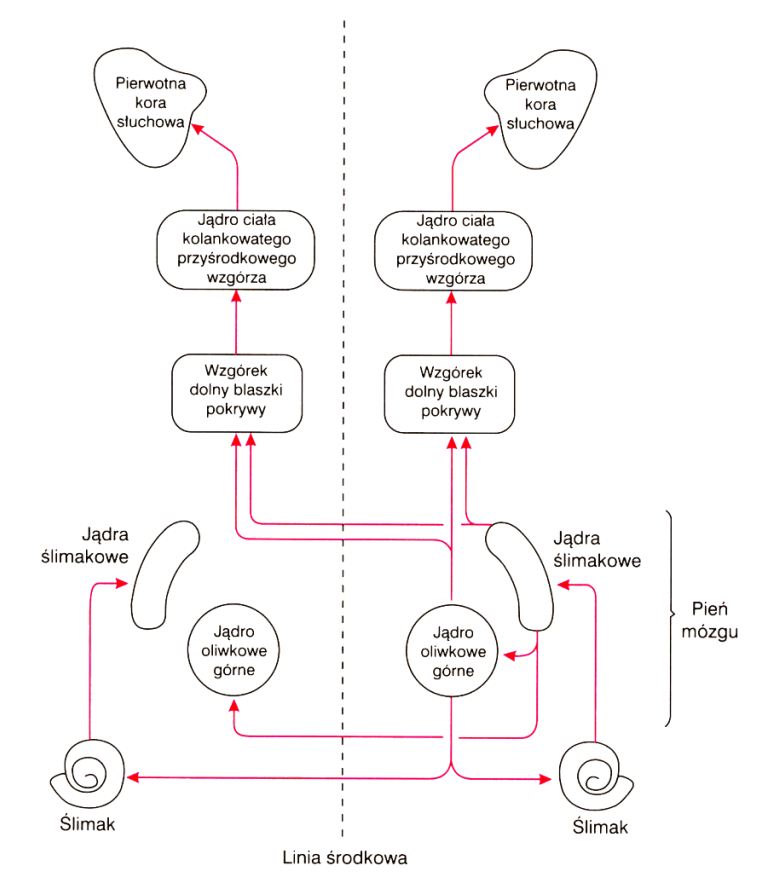
Połączenia po lewej stronie są takie jak po prawej. |
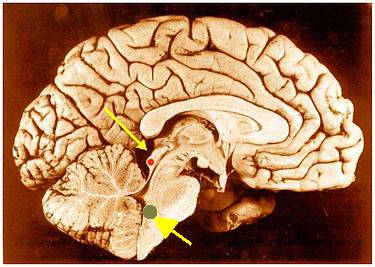
i położenie jąder oliwek. |
Jądra oliwek górnych są niewielkie. Wyróżnia się w nich trzy części, największe to jądro centralne, składające się z około 15 500 neuronów. Górne jądra oliwek przesyłają informację do wzgórków czworaczych dolnych w śródmózgowiu. Tu następuje integracja informacji wzrokowych (wzgórki górne), słuchowych i czuciowych pozwalająca określić istotność doznawanych bodźców i ustalająca napięcie mięśni sterujących ruchem gałki ocznej. Do tej struktury dociera też informacja z kory mózgu. Jak widać kompromis jakim jest ostre widzenie w niewielkim obszarze i widzenie peryferyjne szeroko ale niezbyt wyraźnie wymusił skomplikowany system sterowania sakadami.
|
Sygnał z jąder oliwek wędruje drugą drogą, przez śródmózgowie gdzie łączy się z sygnałami z jąder ślimakowych, do wzgórza, a dokładniej jego części nazywanej jądrami ciała kolankowatego przyśrodkowego (MGN, medial geniculate nucleus), czyli jądra słuchowego wzgórza. Z tego poziomu pętle wzgórzowo-korowe aksonów przesyłają informację pomiędzy wzgórzem i korą słuchową.
Komunikacja zachodzi w obie strony, więc kora może zahamować przesyłanie informacji. Rzadkie przypadki ślepoty lub głuchoty histerycznej mogą być związane z mechanizmem hamowania pobudzeń MGN. Można naprawdę "wytężyć słuch", oczekując dźwięku z określonego kierunku, lub przestać go zauważać jeśli trzeba się skupić na informacjach z innych zmysłów.
Psychiatria zna wiele zaburzeń tego typu, określanych jako
zaburzenia konwersyjne lub dysocjacyjne, a dawniej jako głuchota lub ślepota histeryczna. Uważa się wówczas, że zaburzenia spowodowane są przez czynniki psychologiczne, bo nie widać przyczyn fizycznych, np. uszkodzeń mózgu.
|
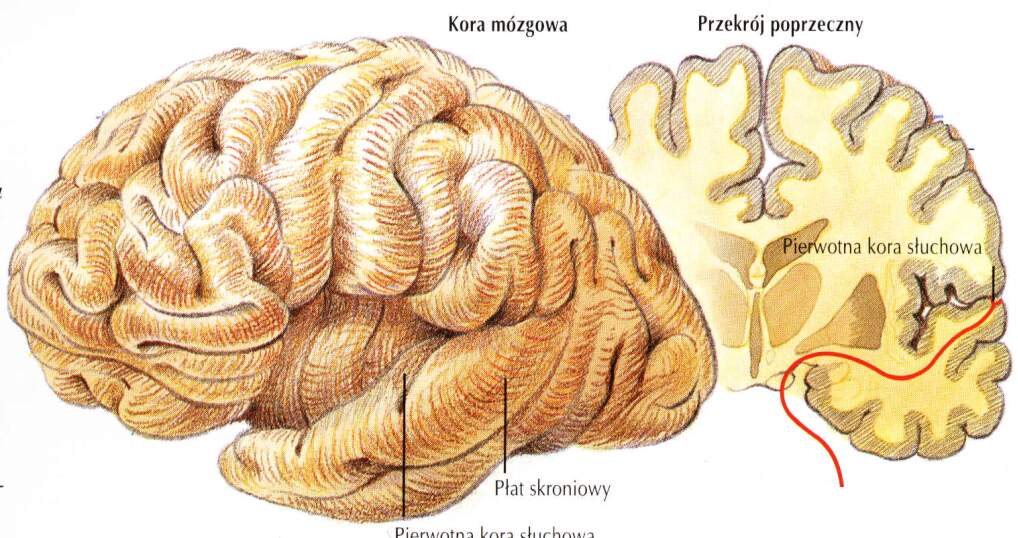
Aktywność pierwotnej kory słuchowej (A1) jest konieczna by powstały świadome wrażenia. Ta kora mieści się w górnym zakręcie skroniowym (STS, superior temporal gyrus). Kora słuchowa zajmuję około 8% całej powierzchni kory. Jej zadaniem jest ocena podstawowych parametrów dźwięku: głośności i wysokości. Zajmuje pola 41/42 Brodmanna, oraz zakręt skroniowy przedni poprzeczny (zakręt Heschla) schowany w bruździe bocznej, pod stykiem płata czołowego i ciemieniowego, na płacie skroniowym.
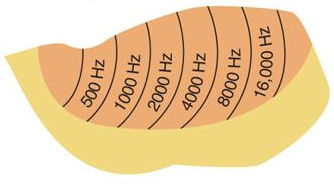
Pierwotna kora słuchowa ma budowę kolumnową. Obserwowane są reakcje na te same częstości tylko przy bardzo słabych, progowych dźwiękach. Projekcja dźwięków na aktywność kory słuchowej jest tonotopiczna, czyli zachowująca podobne aktywacje kory przy podobnych dźwiękach, niskie częstości w częściach bocznych, a wysokie w środkowej części kory.
Selektywna uwaga dotycząca słuchu jest słabiej zauważana niż w przypadku percepcji wzrokowej, która zależy od kontroli sakad i rozszerzenia źrenic. Percepcja słuchowa, jak każda percepcja, jest aktywnym procesem, ale nie ma takich możliwości wpływania na selekcję bodźców jak percepcja wzrokowa. Wiele zwierząt polega przede wszystkim na słuchu. Uwaga może modulować, który obszar kory będzie się silniej pobudzać, dzięki czemu możliwa stanie się spektralna selekcja informacji akustycznych. Badania tego zjawiska (Dick i inn, 2017) pokazały, że ludzie mają zdolności do wybierania najlepszego pasma częstotliwości dzięki modulacji kory słuchowej, wpływając na aktywność obszarów tonotopicznie przetwarzających bodźce słuchowe. Jest to np. istotne dla muzyków, którzy muszą synchronizować swoją grę z innymi instrumentami o określonej wysokości dźwięku, pomimo wielu innych dźwięków instrumentów orkiestry.
|
Większość pobudzeń z lewego ucha dochodzi do prawej półkuli, a z prawego do lewej półkuli. Reakcje kory A1 na podstawowe dźwięki mowy, czyli fonemy, są złożone. Zależnie od rodzaju dźwięków są one dalej przetwarzane w różnych obszarach kory.
W szczególności ponieważ sygnał mowy przetwarzany jest głównie w lewej półkuli słuchanie prawym uchem może prowadzić od nieco lepszego rozumienia mowy niż lewym. Warto zrobić taki test słuchając przez słuchawki niewyraźnej mowy, lub mowy w języku obcym.
Informacje o dźwiękach mózg może też otrzymać dzięki wibracjom rozchodzącym się przez kości. Jest szereg słuchawek, które działają
wykorzystując przewodnictwo kostne. Przyjadą się szczególnie osobom cierpiącym na niedosłuch, ale nie nadają się dla audiofili.
Podsumowując: drogi słuchowe prowadzą od
ślimak ucha wewnętrznego =>
jądra ślimaka =>
jądra oliwek =>
ciało kolankowate przyśrodkowe (MGN, wzgórze) =>
promienistość słuchowa =>
pierwotna kora słuchowa.
Drogi słuchowe
(film, z Digital Anatomist Project).
Uszkodzenia kory powodują osłabienie słuchu w przeciwległym uchu, trudności z lokalizacją dźwięku.
Całkowite uszkodzenie pierwotnej kory słuchowej prowadzi do głuchoty korowej.
Drażnienie prądem kory słuchowej daje wrażenia buczenia, stukania lub brzęczenia
(szumy uszna, określane jako tinnitus).
Czy zwierzęta słyszą ludzką mowę, czy tylko reagują na dźwięki istotne dla ich ekologicznej niszy?
Układ słuchowy jest w znacznej mierze wyspecjalizowany, zwierzęta są wrażliwe na określony rodzaj dźwięków, które mogą być ostrzeżeniem lub przywołaniem. W korze słuchowej mogą istnieć filtry specyficznych sygnałów dźwiękowych, np. ultradźwiękowych pisków, które wydają szczury widząc drapieżnika, lub śpiewu ptaków, które nie zwracają uwagi na inne gatunki, tylko słyszą swoje nawoływania.
W publikacji Gnanateja i inn (2021) odkryto podobieństwa między sygnałem mierzonym bezpośrednio w korze słuchowej (zapisując potencjały elektryczne wewnątrz czaszki) jak i sygnałów EEG ze skóry głowy mózgach ludzi, makaków oraz świnek morskich. Kora we wszystkich przypadkach reaguje podobnie na dźwięki mowy, odtwarzając częstotliwość sygnału akustycznego (frequency following responses, FFRs). Do stymulacja użyto chińskiej sylaby o czterech różnych tonach wznoszenia i opadania, której podstawowe częstotliwości mieściły się w od 89 do 149 Hz.
Reakcje kory różnią się od sygnałów, które pochodzą z pnia mózgu i śródmózgowia. Główny wkład do sygnału na poziomie kory słuchowej pochodzi od pętli wzgórzowo-korowych. Wzgórze przesyła sygnały akustyczne do warstw 3 i 4 kory, a oscylacje w warstwie 3 są zgodne z zmieniającymi się w czasie sygnałami mowy, są więc reprezentowane na poziomie korowym.
|
Wtórna kora słuchowa: wokół pierwotnej kory
obszar Wernickego w płacie skroniowym półkuli dominującej .
Lezje wtórnej kory słuchowej wywołują różnorodne
afazje (gr. a+phasis, brak mowy).
- W dominującej (zwykle lewej) półkuli: blisko kory pierwotnej, górny zakręt skroniowy => zaburzenia słuchu fonematycznego, trudności w rozróżnianiu wyrazów (agnozja słuchowa).
- W przeciwległej (zwykle prawej) półkuli: słabsza percepcja melodii i rytmu,.
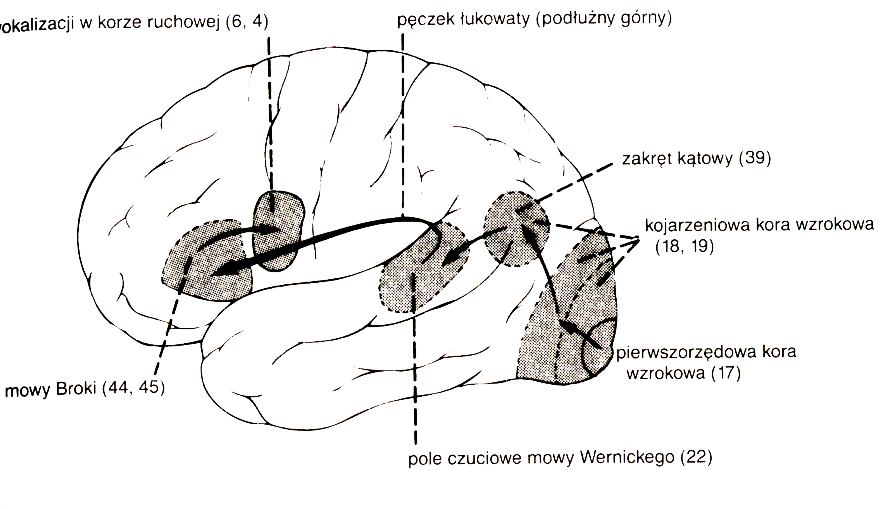
Połączenia międzykorowe:
- wtórne obszary słuchowe w korze skroniowej; w dominującej półkuli ośrodek Wernickego w tylnej części górnego zakrętu skroniowego, związany ze rozumieniem mowy;
- ośrodek Broki związany z produkcją mowy w tylnej części płata czołowego;
- kora słuchowa z przeciwległej półkuli (lokalizacja przestrzenna);
- kora czuciowa (dźwięki wywołują reakcje czuciowe nie tylko przez drgania!);
- FEF (Frontal Eye Fields), czołowy ośrodek okoruchowy (świadoma orientacja oczu).
|
Wczesna specjalizacja kory przy rozróżnianiu fonemów prowadzi do trudności w uczeniu się języków obcych, np. języków tonalnych. Polakom trudności sprawia rozróżnienie np. angielskiego "thieve" i "sieve", japończykom jednakowe wydaje się "ram" i "lam".
Zdolność do rozróżniania fonemów języków obcych zanika ok. 10 miesiąca życia; jest to przyczyną trudności uczenia się języków obcych w starszym wieku: jeśli się wyraźnie nie słyszy to nie można też nauczyć się rozumieć i mówić.
Agnozja słuchowa prowadzi do upośledzenia mowy (brak rozumienia potrzeby korekcji, skoro się nie słyszy różnicy).
Rozwój map tonotopicznych jest jedną z podstaw dobrego słuchu.
Hałas, biały szum, powoduje brak zróżnicowania map w korze słuchowej u szczurów i zapewne u ludzi; sprzyja to wywoływnaiu ataków padaczki.
Niemowlęta dorastające w hałasie mogą mieć trudności językowe, a być może nawet zaburzenia podobne do autyzmu: ich mózgi muszą się nauczyć filtrować informację i dłużej skupiać wyłącznie na jednym bodźcu, co może spowodować trudności z przenoszeniem uwagi.
Kontrastowe, proste bodźce w ciszy najlepiej rozwiną mapy słuchowe.
Wiele osób cierpi na szumy uszne (tinnitus), powstające na skutek spontanicznych pobudzeń nerwów słuchowych. Takie "fantomowe" dźwięki nie muszą być szumami, może to być pisk, gwizd, ćwierkanie, tykanie czy dzwonienie w uszach. To dość częste schorzenie, może być na tyle uciążliwe, by wymagać leczenia.
|
Afazja Broca (eferentna afazja ruchowa) jest wynikiem uszkodzenia obszaru Broca i prowadzi do trudności z wymówieniem słów, ciągów sylab, czasami pozostaje tylko kilka słów, które da się wymówić.
Afazja Wernickiego (akustyczno-gnostyczna afazja skroniowa), powstaje po uszkodzeniu obszaru Wernickiego i obszarów przyległych, prowadząc do zaburzeń rozumienia mowy własnej i innych.
Mowa jest płynna, ale niegramatyczna, bez sensu, słowa są pomieszane, tracą znaczenie.
Niezdolność rozumienia mowy prowadzi do niezdolności do pisania i czytania.
Pacjenci są nieświadomi swoich problemów językowych! To ładny przykład problemu, z którym zmierzyć się musi teoria świadomości - jak to możliwe, by słysząc siebie nie zauważyć, że mówimy bez sensu?
Uszkodzenie samego obszaru Wernickiego powoduje czystą głuchotę słów (Kirshner, 2006)
Afazja przewodnictwa wynika z uszkodzenia pęczka łukowatego, łączącego ośrodek Wernickiego i Broca.
Mowa pozostaje płynna i rozumienie normalne, ale są trudności w powtórzeniu słowa i częste przekręcanie wyrazów, pomimo chęci skorygowania błędów.
|
Lezje zakrętu skroniowego środkowego dają zaburzenia pamięci słuchowo-werbalnej
(afazja akustyczno-mnestyczna).
Efekt: niezdolność do powtórzenia serii wyrazów.
Lezje tylnej części dominującego płata skroniowego:
afazje nazewnicze.
Powodem jest brak skojarzeń między wzrokowym rozpoznaniem a nazwą.
Nie można narysować przedmiotu po usłyszeniu instrukcji słownej, chociaż nie ma trudności w kopiowaniu rysunków.
Zagadnienia związane z mową związane są też z wyższymi czynnościami psychicznymi i zostaną omówione później.
Afazja akustyczna: zaburzenie rozpoznawania dźwięków niewerbalnych, odgłosów zwierząt lub sygnałów akustycznych (używana jest też nazwa
"agnozja akustyczna").
Testy: wskazać obrazek kojarzący się ze źródłem dźwięku.
Dwa rodzaje afazji akustycznej: niezdolność do odróżniania dźwięków (kora pierwotna) i niezdolność do ich kojarzenia (kora wtórna).
Afazja dwujęzyczna (bilingual aphasia) pokazuje, że można zapomnieć selektywnie rozumienia jednego języka a jednocześnie w nim mówić; zdarza się też niekontrolowane mieszanie języków.
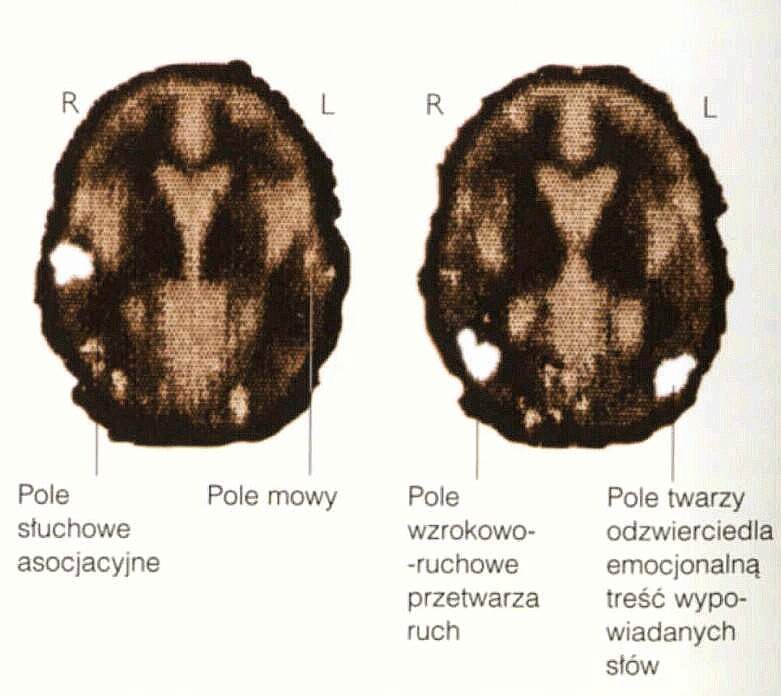
|
Interpretacja emocjonalna wrażeń słuchowych jest silnie uwarunkowana kulturowo, np. siorbanie i siąkanie Japończyków i Chińczyków wywołuje u nas nieprzyjemne reakcje, a nasze głośne smarkanie w chusteczkę podobne reakcje u nich.
Bicie serca może mieć efekt kojący dla niemowląt, ale czy naprawdę "kołysanki prenatalne" mają na dorosłych pozytywny wpływ? Trudno tu znaleźć wiarygodne wyniki badań.
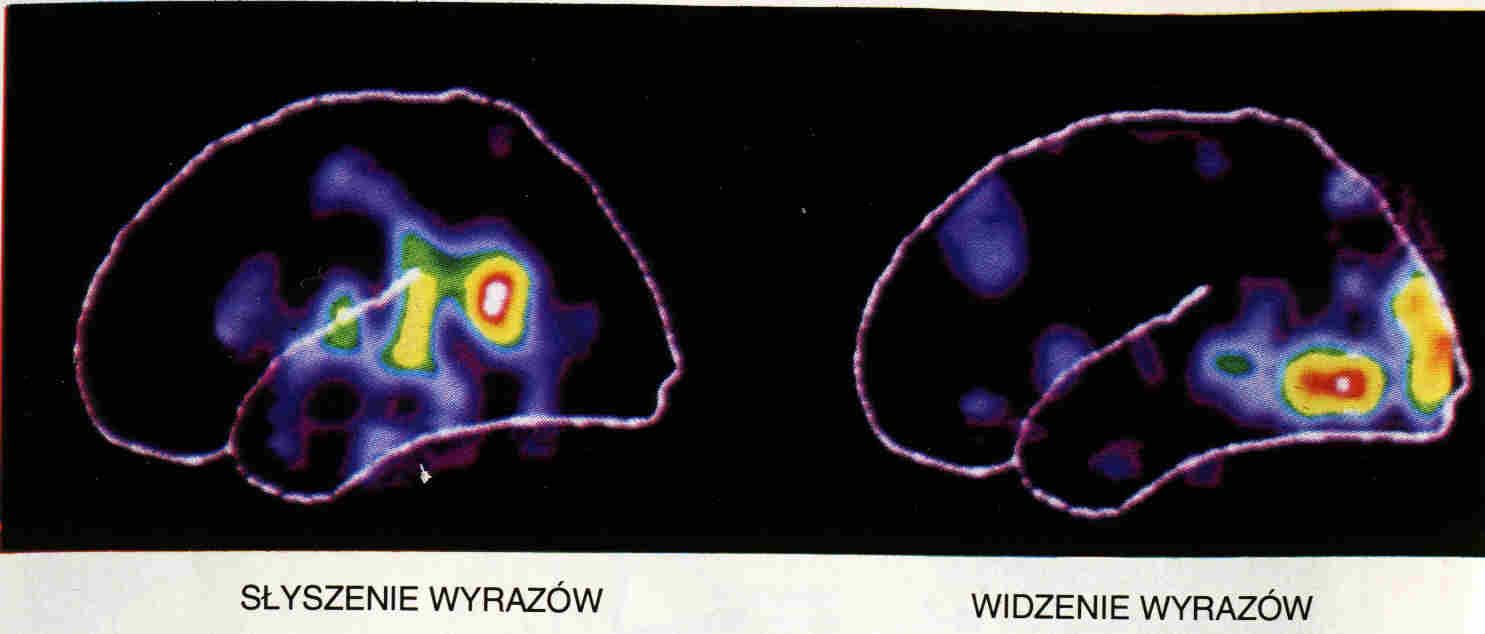
Możliwe jest "słuchanie" na podstawie obserwacji
ruchów warg.
Można się tego nauczyć dzięki
komputerowym aplikacjom.
Układ warg można podzielić na kategorie zwane
wisemami.
Jest wiele wersji języków migowych.
Uniwersytet Gallaudeta w USA istnieje od 1857, naucza się tam korzystając z
Amerykańskiego Języka Migowego (ASL) osoby głuche (Sacks, 1998).
Podobnie jak iluzje wzrokowe, istnieją też
iluzje słuchowe, związane z mową, dźwiękami muzyki, słuchem przestrzennym, oddziaływaniem między wzrokiem a mową.
Jest to obszerny i ciekawy temat, sporo przykładów jest na stronie
Diane Deutsch, która napisała o iluzjach akustycznych kilka książek i
przykłady odkrytych przez nią iluzji.
Can you trust your ears (YouTube) prezentuje efekt McGurka (wpływ wzroku na wrażenia akustyczne) i kilka iluzji.
To popularny efekt, często wykorzystywany w różnych prezentacjach.
"Yanny" or "Laurel"? Wyjaśnienie.
5 akustycznych iluzji (New Scientist) - potrzebne są słuchawki.
Holofonia, czyli nagrania za pomocą sztucznej głowy, odwzorowują przestrzennie z dużym realizmem położenie źródła dźwięków. Jest też wiele systemów tworzenia dźwięku przestrzennego 3D, najlepiej odbieranego za pomocą słuchawek.
Osoby, które straciły słuch mogą doznawać halucynacji słuchowych, ale osoby głuche od urodzenia mają jedynie halucynacje wzrokowe sugerujące mowę (gesty, ruch ust).
Około 5% zdrowych ludzi ma halucynacje słuchowe, ma je za to większość schizofreników (Frontiers of Human Neuroscience 2013, University of Bergen Group).
Percepcja muzyki
Efekt Mozarta, znany z popularnej literatury, to wpływ słuchania muzyki klasycznej (od Vivaldiego i Bacha do Mozarta) na wyników testów.
Wiarygodnie pokazano pozytywny wpływ muzyki na pamięć roboczą i rozumowanie czasoprzestrzenne (Rauscher i inn. 1993); nie oznacza to jednak trwałego zwiększenia inteligencji, a raczej chwilowe pobudzenie kory i polepszenie nastroju.
Nawet biały szum w słuchawkach wpływa na chwilowo lepsze wyniki w zadaniach arytmetycznych.
Nie chodzi o Mozarta tylko o muzykę pobudzającą i podnoszącą nastrój, efekt jest dość słaby, ale w pewnych warunkach zauważalny; prasa i firmy komercyjne bardzo go wyolbrzymiły.
W stanach Tennessee i Georgia każdy nowo narodzony obywatel dostaje CD z muzyką Mozarta - krytycy
kwestionują jednak, czy jest to program sensowny.
Efekty są kontrowersyjne, różnią się u ludzi starszych, dorosłych i u dzieci, zależą też od rodzaju muzyki, prawdopodobnie wynika to z indywidualnych upodobań, ale IQ od tego nie wzrasta.
Mozarta oczywiście warto słuchać, a jeszcze bardziej grać samemu, wtedy
efekty będą wyraźniejsze.
Zbadano efekty muzyki na osoby cierpiące na różne schorzenia, np. padaczkę, ale "efekt Mozarta" to głównie
efekt medialny a nie rzeczywisty.
Muzykoterapia ma liczne zastosowania, muzyka relaksacyjna może np. spowodować obniżenie ciśnienia i częstości napadów padaczki, stosowana jest w rehabilitacji poudarowej, stanów nerwicowych ("muzyka koi nerwy") i innych problemach.
Meloterapia to muzykoterapia śpiewem.
Choreoterapia to terapia tańcem.
Doświadczenia na szczurach pokazały pozytywny wpływ na rozwój i sprawność działania ich mózgów.
Słuch absolutny jest rzadki, ma go 1 osoba na 10.000.
W kulturach, które posługują się językiem tonalnym (np. chińskim czy wietnamskim) występuje znacznie częściej (por. Diana Deutsch).
Amuzja to niezdolność do określenia względnej wysokości dźwięku, powtórzenia melodii;
zwykle prowadzi to do całkowitego braku zainteresowania muzyką.
Amuzja prawie nie występuje tam, gdzie używane są języki tonalne.
Śpiewanie ma dobroczynny wpływ na osoby z chorobą Alzheimera, Parkinsona i inne, pobudzając emocje.
Śpiewanie czy wybijanie rytmu ma wpływ na synchronizację procesów w mózgu.
(How singing unlocks the brain).
Biologicznymi wpływami muzyki zajmuje się biomuzykologia; można w niej wyróżnić:
- muzykologię kognitywną (nazywaną też neuromuzykologią);
- etnomuzykologie, czyli etnografię muzyczną;
- muzykologię ewolucyjną, zajmującą się przyczynami rozwoju zdolności muzycznych.
Strojenie instrumentów i skale muzyczne.
Dwa dźwięki, których częstotliwości podstawowe są ze zgodne postrzegane są jako harmonijne, czyli brzmią unisono. Dzięki przyjemnie brzmiące, czyli
konsonanse, różnią się od siebie
interwałami.
Dźwięki o dwa razy wyższej częstotliwości brzmią podobnie, gdyż oprócz dźwięku podstawowego powstają zwykle dźwięki harmoniczne (np. pobudzenie pierwszej struny E w gitarze wywołuje wibracje szóstej struny E).
Jak można podzielić przestrzeń dźwięków tak, by brzmiały przyjemnie? Stosunek wysokości dźwięków powinien być naturalnym ułamkiem. Oktawa to stosunek częstotliwości 2:1, np. A'=440 Hz i A''=880 Hz, te dwa dźwięki słyszane jednocześnie brzmią przyjemnie bo nie ma między nimi dudnień. Naturalna skala dźwięków może wybrać kilka o wysokościach, których częstotliwości f'/f są naturalnym ułamkiem, np. 5 dźwięków skali pentatonicznej (używanej w muzyce ludowej, w tym blusie), skali diatonicznej (7 dźwięków, białe klawisze) lub skali chromatycznej, dodającej do 7 dźwięków jeszcze 5 (czarne klawisze). Różne tonacje wybierają te 7 dźwięków tak, by jak najlepiej ze sobą harmonizowały.
Jeśli instrumenty mają ze sobą dobrze współbrzmieć trzeba ustalić sposób ich nastrojenia. Skala złożona z 12 dźwięków jeszcze za czasów Bacha nie była uniwersalna, ale już w 17 wieku zaczęto dzielić oktawę w taki sposób, że stosunek wysokości dwóch kolejnych dźwięków, różniących się o pół tonu, to
12√(2). Taki system nazywa się
równomiernie temperowanym i jest obecnie powszechnie stosowany w muzyce zachodniej.
To znacznie uprościło tworzenie muzyki dla wielu instrumentów, ale wiele osób uważa, że lepszą podstawą dla skal muzycznych powinna być częstotliwość 432 Hz lub inne częstotliwości. Jest na ten temat
wiele informacji i wideo.
Większość ludzi nie ma słuchu absolutnego i postrzeganie wysokości dźwięków nie jest doskonałe. Dlatego można było zdefiniować podział równomiernie temperowany, chociaż nie wszystkie dźwięki tworzą w nim doskonałe konsonanse. Dudnienia są niewielkie, więc można wybrać dźwięki tak, by nie było słychać dysonansów. Najmniejsze odchylenia od naturalnego stroju są dla kwarty (5 półtonów) i kwinty (7 półtonów), czyli np. kwarty A'-D" naturalny stosunek to 4:3=1.333, czyli 440*4/3=586.666 Hz, a temperowany 587.330, a dla kwinty A'-E" to 3:2, czyli 660 Hz, a temperowany to 659.255 Hz. Takie różnice są dla większości osób niesłyszalne, ale są też konsonanse niedoskonałe, np. 3 półtony, A'-C'' gdzie zamiast ilorazu 6:5 (528/440 Hz) mamy 523.16 Hz, a więc ok. 4 Hz różnicy.
Wśród muzyków można spotkać opinię, że strojenie do A'=432 Hz zamiast 440 Hz ma większy pozytywny wpływ na ludzi. Na razie jedyne badanie pilotażowe
(Calamasi, Pomponi 2019)
na stosunkowo niewielkiej grupie 33 osób pokazało niewielkie zmiany, spowolnienie pulsu i oddechów, ale wymaga to dalszych badań.

|
Jaka była wartość ewolucyjna muzyki?
Dlaczego słuchanie muzyki sprawia nam przyjemność?
Mowa konieczna jest do komunikacji. Do czego potrzebna była muzyka?
Steven Pinker: "Jeśli chodzi o biologiczne przyczyny i efekty muzyka jest bezużyteczna".
Czyżby muzyka nie miała ewolucyjnego sensu? Pinker pisał to w odniesieniu do bardzo wczesnego rozwoju Homo sapiens, nie należy więc sobie wyobrażać ludzi współczesnych, dla których muzyka ma oczywiste znaczenie.
Śpiew samców ptaków, wielorybów, czy zawodzenia gibbonów służą przywabianiu samic, kojarzone z przyjemnością, próbą przyciągnięcia uwagi. Może też być wyrazem smutku, pokazującym potrzebę współczucia.
Już w starożytnych Chinach pisano:
Smutne wołania gibonów w trzech kanionach Pa-Tu,
Po trzecim ich koncercie tej nocy,
mokre od łez jest ubranie wędrowca.
Poemat Chiński z 4 wieku.
B. Merker (1999), biomuzykolog: wokalizacja małp stadnych służy komunikacji i ostrzeganiu obcych małp, jest tańsza niż ślady zapachowe.
Ksenofobia prowadzi do dziedzicznych chorób; potrzebne jest mieszanie genów.
Potrzebny jest sygnał pozwalający przekroczyć barierę strachu i przejść do innej grupy.
U szympansów (i ludów myśliwsko-zbierackich) samice wabione są głosem chóru samców.
Istniała ewolucyjna presja do wspólnej wokalizy.
Na poziomie psychologicznym związek śpiewu z szukaniem partnera, miłością i seksem jest wyraźny.
Na poziomie neurofizjologicznym śpiew pobudza ośrodki przyjemności, wyzwala mechanizm nagrody za przezwyciężenie barier między osobnikami.
Na poziomie społecznym współpraca samców zwiększała spójność wewnątrz grupy.
Piszczałki i bębny są słyszalne na większe odległości, można je używać do sygnalizacji
("mówiące" bębny tama w Afryce), były więc powody do używania instrumentów.
Jest wiele innych hipotez tego rodzaju rozwiniętych w ramach
ewolucyjnej muzykologii.
Współczesna muzyka klasyczna tworzy złożone, bogate struktury dźwięków.
Co w nich nas porusza? Czy są to te elementy, które do jakiegoś stopnia się nakrywają z naturalnymi dźwiękami mającymi znaczenie ewolucyjne? Być może nie rozumiemy jeszcze istoty oddziaływania muzyki.
Wpływ muzyki na abstrakcyjne myślenie zbadany został na przykładzie uczenia się ułamków w trzeciej klasie.
Znajomość nut, wyczucie związanego z tym rytmu, ułatwia uczenie się ułamków.
Cymatyka
pozwala zobaczyć dźwięki, wibracje w mózgu powstające w wyniku dochodzących sygnałów są dużo bardziej skomplikowane i zachodzą w trzech wymiarach, chociaż lepiej jest je analizować w przestrzeni o większej liczbie wymiarów, np. aktywacji różnych podsieci.
CymaScope i
CymaPiano
i cymatyczna próba zrozumienia
mowy delfinów za pomocą "dźwiękowych obrazków".
Do tematu percepcji muzyki będzie przy omawianiu złożonych czynności mózgu.
Rozważaliśmy tu mechanizmy przetwarzania informacji dźwiękowej.
Filozofia percepcji słuchowej szuka odpowiedzi na wiele pytań: czym właściwie są dźwięki, czy są prywatnymi odczuciami czy publicznym zdarzeniami? Takie pytania dotyczą sposobu interpretacji na poziomie zdarzeń mentalnych.
B16.3 Zmysł równowagi i orientacja w przestrzeni
|
Tylna część kory wyspy i część
płata ciemieniowego otrzymuje sygnały z układu zachowania równowagi.
Stymulacja tego obszaru wywołuje wrażenia ruchu i obrotów.
Pobudzenia padaczkowe tego obszaru wywołują też takie wrażenia.
Uszkodzenia wywołują zanik zdolności do dyskryminacji ruchu.
Wzrok może dostarczać wystarczającej informacji dla orientacji.
W ciemności, silnym deszczu lub śniegu to jednak nie wystarcza.
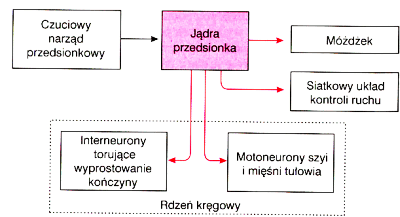
|
Układu równowagi wraz z układem czuciowym, wzrokiem i priopriocepcją bierze udział w tworzeniu mapy przestrzeni wokół ciała (peripersonalnej).
Przestrzeń bliska, w której możemy działać, jest inaczej reprezentowana niż przestrzeń odległa.
Zaburzenia reprezentacji przestrzeni peripersonalnej wywołują ciekawe efekty:
Obraz świata i relacji przestrzennych jest kombinacją egocentrycznego i allocentrycznego punktu widzenia.
Allocentryczne (od greckiego słowa "inny") postrzeganie, związane z obiektami, rozwija się pomiędzy 3 a 5 rokiem życia, kiedy dziecko zaczyna sobie wyobrażać świat z innej perspektywy niż tylko egocentryczna.
Allocentryczne postrzeganie jest skupione na obiektach, ich relacjach, własnościach, ich formie niezależnie od naszego ruchu i położenia, jest w znacznej mierze niezależne od położenia obserwatora - rozpoznajemy obiekty i znamy ich własności, chociaż im się szczegółowo nie przyglądamy, wiążemy je z pewną kategorią.
Mechanizmy przetwarzania informacji ego i allocentrycznej są częściowo znane dzięki pracom zrobiony w większości już w 21 wieku.
Allocentryczne postrzeganie wiąże się z brzusznym szlakiem wzrokowym, rozpoznawaniem obiektów jako takich, bez odwołania do swojego położenia.
Egocentryczne postrzeganie wiąże się z aktywnością prawego górnego płacika ciemieniowego
(SPL), analizującego informację o tym, gdzie są w danej chwili nasze własne kończyny i jakie są ich możliwości działania uwzględniając wielkość postrzeganego obiektu.
Okolice tylnej bruzdy ciemieniowej są połączone z płatem czołowym i pełnia rolę w kontroli sakad, uwadze wzrokowej, działaniach wolicjonalnych związanych z wskazywaniem i chwytaniem.
Grzbietowy szlak wzrokowy odpowiada więc na pytanie "gdzie to jest w relacji do mnie i co mogę z tym zrobić", a nie tylko na pytanie ogólne "gdzie". Jest to konieczne by określić, czy mogę dany obiekt złapać ręką lub dosięgnąć nogą, czy warto próbować, jak ocenić głębię, siłę i miejsce uderzenia?

|
Stymulacje elektryczne prawej półkuli w okolicach zakrętu kątowego mogą wywołać różne wrażenia autoskopowe:
- oddzielenie od własnego ciała (out-of-body experience);
- zmianę perspektywy widzenia, np. unoszenie się pod sufitem, zachowując perspektywę egocentryczną;
- wrażenie widzenia swojego ciała z perspektywy zewnętrznej;
- negatywna autoskopia, w której nie widzi się swojego odbicia w lustrze;
- wewnętrzna autoskopia: widzi się wewnętrzne organy ciała patrząc na swoje odbicie w lustrze.
Przyczyny powodujące takie wrażenia to lezje, wpływ substancji halucynogennych, wyczerpanie organizmu (np. śmierć kliniczna), lub stany podobne do ataków padaczki, które mogą spowodować chwilowe zaburzenia pracy mózgu.
W przeszłości (i nadal) takie halucynacje uznawano za dowód istnienia umysłu niezależnego od ciała ...
Sugestie hipnotyczne mogą również zaburzyć mechanizm reprezentacji przestrzeni i wrażenie wolicjonalnego działania, wywołując zespół "obcej ręki".
Ruchy stolika (ouija board) lub talerza, na którym trzyma ręce grupa osób w czasie seansu spirytystycznego też należą do tej kategorii - uczestnicy są przekonani, że nie popychają stolika. Michel Faraday w 1853 roku zrobił prosty eksperyment pokazujący, że to nie stolik ciągnie ręce tylko ręce pchają stolik, chociaż mamy całkiem przeciwne wrażenie.
Złudzenia ideomotoryczne potrafią być bardzo silne:
Science of Scams ma różne wideo pokazujące jak robią to spirytyści i wróżbici.

|
Inne zaburzenia relacji przestrzennych:
- Allotopagnozja: ogólna niezdolność do wskazywania rzeczy.
- Autotopagnozja (Pick, 1922) to niezdolność do wskazywania własnych części ciała po otrzymaniu werbalnego polecenia.
- Heterotopagnozja: niezdolność do wskazywanie części ciała innych osób.
Autotopagnozja wiąże się z lezjami lewego płata ciemieniowego, pacjenci nie mają problemu z wskazywaniem i dotykaniem np. części maszyn;
nie mogą też narysować czy prawidłowo poskładać układankę przedstawiającą części ciała (inna nazwa to somatotopagnozja, Gerstmann 1942), podkreślająca, że jest to niezdolność do prawidłowego postrzegania ciała.
Autotopagnozja często występuje razem z heterotopagnozją, ale znane są nieliczne przypadki czystej heterotopagnozji (lezje górnej kory ciemieniowej) i czystej autopagnozji (lezje dolnej kory ciemieniowej).
Pacjenci z heterotopagnozją nie potrafią wskazać (funkcja komunikacyjna, wskazywanie na kogoś), a potrafią uchwycić (działanie bez komunikacji).
mogą też w niektórych przypadkach wskazywać części ciała innych osób zamiast swoich (Langavant i inn, 2009) oraz dotknąć odpowiedniej części, a nie wskazać. Są to rzadkie przypadki i nie ma tu jednoznacznego opisu.
Są też liczne inne zaburzenia związane z zaburzeniem postrzegania ciała, np. autoprosopagnozja, niezdolność do rozpoznawania własnej twarzy (Vignemont 2009).
B16.4 Smak
Pozostałe zmysły również potrzebują kory mózgu do precyzyjnej dyskryminacji wrażeń.
|
Adaptacja do smaku: receptory przestają reagować, pomimo ciągłego pobudzania. Podobnie jak iluzja wodospadu w przypadku wzroku również nasze receptory smaku i zapachu w krótkim czasie ustalają nowy stan odniesienia i reagują na zmiany a nie stan ustalony. Zjedzenie góry lodów nie daje wiele przyjemności tylko ból brzucha, zjedzenie małej gałki skupiając się na wrażeniach smakowych może sprawić dużo przyjemności.
Wzajemne wpływy, np. adaptacja do kwasu (np. kwasku cytrynowego) może wywołać wrażenie słodkości wody.
Fenylotiokarbamid (PTC) przez 65% populacji uważany jest za mocno gorzki, a 35% nie czuje wcale smaku - odpowiedzialna jest za to pojedyncza mutacja genetyczna.
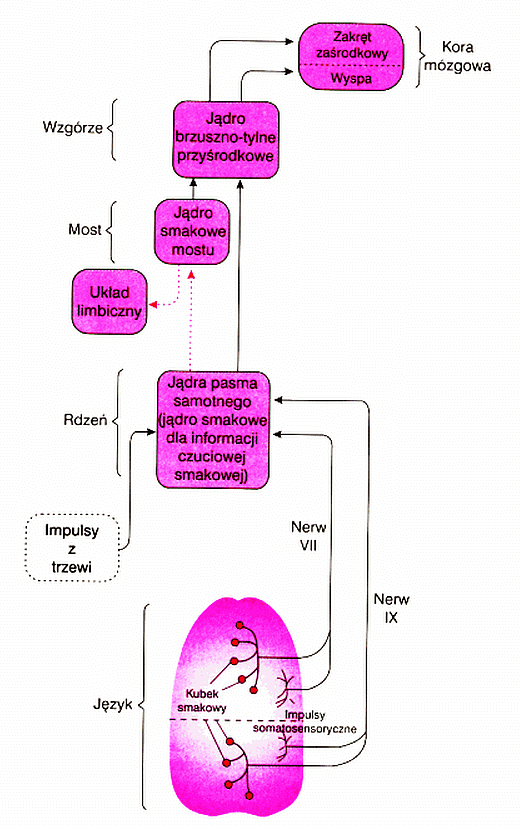
Informacja na temat aktywności kubków smakowych wędruje od chemoreceptorów wiązką nerwów VII i IX przez rdzeń kręgowych do znajdującego się w pniu mózgu jądra pasma samotnego, a następnie przez jądro smakowe mostu rozgałęzia się, trafiając do struktur układu limbicznego, gdzie analizowana jest przyjemność lub powstają nieprzyjemne reakcje związane ze smakiem.
Druga droga prowadzi przez tylno-brzuszno-przyśrodkowe jądro wzgórza do kory smakowej i kory wyspy, która bierze udział w reakcjach awersyjnych, w tym wstrętu.
Kora smakowa mieści się w zagłębieniu płata ciemieniowego, okolicach zakrętu zaśrodkowego, niedaleko kory somatosensorycznej SI reprezentującej język. Jej uszkodzenia powodują zanik zdolności rozróżniania smaków.
|
Receptory w kubkach językowych przesyłają informację przez jądra pasma samotnego znajdujące się w pniu mózgu. Te jądra łączą się z jądrami smakowymi w moście, oraz bezpośrednio ze wzgórzem. Jądra smakowe mostu przesyłają sygnały do układu limbicznego, jak i do wzgórza.
Pobudzenia kory smakowej (np. padaczkowe) w wyniku wewnętrznych procesów w mózgu wywołują halucynacje smakowe, czasami to oznaki nadchodzącego ataku padaczki.
Halucynacje różnego rodzaju są dość częste (zdarzają się ponad 1/4 ludzi na jawie), a węchowe i smakowe halucynacje są najczęstsze.
Zespół smakosza (Gourmand syndrome): uszkodzenie prawego płata czołowego, związanego z układem nagrody, może wywołać obsesję na punkcie wykwintnego jedzenia.
Smak zmienia się w zależności od potrzeb organizmu:
brak witamin czy soli spowoduje po dłuższym czasie (np. dryfowaniu rozbitka na morzu) zmiany smaku tak, że chętnie je się wnętrzności i oczy ryb.
Pieprz syczuański wywołuje ciekawą reakcję, przypominającą wibracje na języku; mechanizm tej reakcji został
odkryty w 2008 roku.
B16.5 Węch
Ludzie mają około 40 mln komórek węchowych w obszarze około 2 na 5 cm; psy około 1 mld, a najlepszy węch mają niedźwiedzie (oceniany na 7 razy lepszy niż psy).
Znamy około 1000 różnych
receptorów węchowych,
ale każdy zapach pobudza wiele z nich i każdy receptor reaguje na wiele zapachów.
Kodowanie zapachów ma kombinatorycznych charakter, podobnie jak słowa zapisywane za pomocą liter różne wrażenia zapachowe związane są z pobudzaniem kombinacji receptorów (mechanizm ten częściowo poznano w 1999 roku).
U człowieka ponad 400 genów koduje białka receptorów węchowych.
Kobiety mają średnio lepszy węch, ale są duże indywidualne różnice. Ewolucyjnie wydaje się, że Homo Sapiens miał nieco lepszy węch niż wymarłe gatunki hominidów.
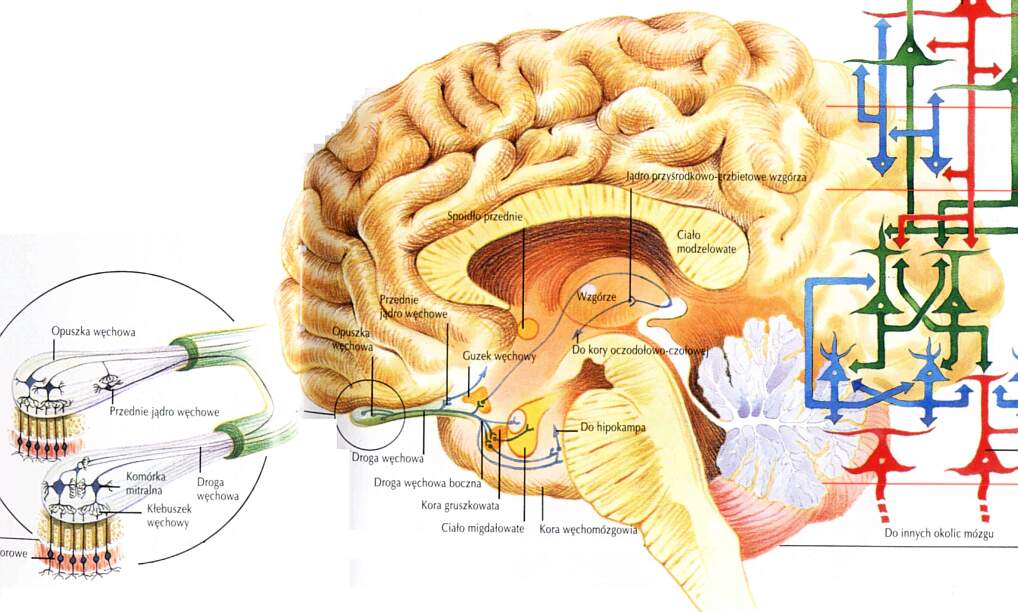
Ok. 5 mln neuronów przesyła sygnały przez nerw węchowy do skupisk neuronów w opuszce węchowej, zewnętrznym, pierwotnym fragmencie kory (neurony opuszki podobne są do komórek nabłonka skórnego).
Węch doprowadza informacje bezpośrednio do ciała migdałowatego układu limbicznego => szybkie reakcje emocjonalne, strach wywołany zapachem.
Uszkodzenia kory węchowej powodują nieprzyjemne halucynacje węchowe, czasami to oznaki ataku padaczki.
Drogi pobudzeń (Kolb & Whishaw, 1996):
- Receptory => opuszka węchowa => kora gruszkowata i otaczające ciało migdałowate => boczna część podwzgórza => kora oczodołowo-czołowa.
- Receptory => opuszka węchowa => kora gruszkowata i otaczające ciało migdałowate => jądro przyśrodkowe-grzbietowe wzgórza => kora oczodołowo-czołowa.
- Receptory => opuszka węchowa => kora gruszkowata => kora węchomózgowa => hipokamp.
|
Wtórna kora węchowa:
hak (brzuszna część kory czołowej) i
kora śródwęchowa, bliska formacji hipokampa.
Jedyny system, który nie łączy się z korą wyłącznie przez wzgórze; większość kory węchowej to kora stara, 3-warstwowa, ale kora śródwęchowa ma 6 warstw.
Interpretacja wrażeń węchowych jest bardzo indywidualna, zależna od wspomnień, często niejasna - pobudzenie struktur limbicznych i niespecyficzne pobudzenia różnych okolic kory.
Kora oczodołowa jest obszarem, w którym schodzą się informacje o smaku, zapachu, dotyku i rozpoznaniu wzrokowym, ma silne projekcje do prążkowia i bocznej części podwzgórza.
Zaburzenia smaku i węchu.
- Anosmia to brak wrażeń węchowych, zaburzenie na które cierpi około 5% populacji.
- Hyposmia - zmniejszona wrażliwość węchowa.
- Hyperosmia - zwiększona wrażliwość na niektóre lub wszystkie zapachy.
- Dysosmia - ogólne zmiany percepcji węchowej:
- Kakosmia, to napady wrażeń, że odczuwa się wstrętne zapachy.
- Parosmia - normalne zapachy wydają się odrażające;
- Phantosmia, czyli fantomowe wrażenia węchowe, halucynacje węchowe.
- Heterosmia - zanik rozróżniania niektórych zapachów.
Analogicznie z zaburzeniami smaku:
- Ageuzja,
- hypogeuzja,
- dysgeuzja, i hypergeuzja to brak, redukcja, zniekształcenie i zwiększona wrażliwość smakowa.
Wrodzona anosmia. skorelowana jest z wrodzoną analgezją (brakiem odczuwania bólu), dochodzi do niej z powodu mutacji genu SCN9A, którego białka tworzą sodowy kanał jonowy Nav1.7 w receptorach bólowych (nocyceptorach) jak i w receptorach węchowych.
Wszyscy w znacznym stopniu cierpimy na anomię zapachów - potrafimy rozpoznać a przynajmniej rozróżnić bardzo wiele zapachów, ale nazwać tylko nieliczne z nich, większość uznając za przyjemnie pachnące bądź śmierdzące.
W niektórych kulturach zapachy grają większą rolę, a nazywania zapachów jest równie łatwe jak nazywanie kolorów. Przykładem jest lud zbieracko-myśliwski Jahai z Półwyspu Malajskiego
(Majid, Kruspe 2018), który radzi sobie znacznie lepiej niż ludzie z innych grup etnicznych. Jest to związane z ich trybem życie i rolą zapachów.
Przemysł perfumeryjny
zatrudnia osoby o wyjątkowej wrażliwości na zapachy. Są to ludzie, którzy mają wyobraźnię zapachową i potrafią tworzyć kompozycje zapachowe w swojej głowie. Talent twórców perfum zależy od ich wyobraźni: badania fMRI pokazały reakcję kory węchowej pod wpływem wyobraźni. To rzadka umiejętność wymagająca długiego treningu, pod jego wpływem kora węchowa reaguje silniej na zapachy robi się nieco grubsza.
Układ wzrokowy rozdziela sygnały wzrokowe niosące bogatą informację. Rozpoznawanie przedmiotów w oparciu o kolory i kształty, czy zwierząt przy uwzględnieniu ruchu, polega na przypisaniu do jednej kategorii bardzo różnych stymulacji wzrokowych. Wyspecjalizowane obszary kory analizują różne cechy sygnałów wzrokowych niezależnie, dzięki czemu łatwiej jest im przypisać dyskretne cechy pozwalające na kategoryzację wzrokowych bodźców, pomimo znacznej zmienności obrazu dla każdej kategorii. W przypadku zapachu projekcje kory zapachowej docierają do bardzo wielu podkorowych obszarów mózgu (ciało migdałowate, hipokamp, wzgórze, podwzgórze), pobudzając całą korę i przywołując liczne wspomnienia. Pobudzenia kory są więc mało specyficzne.
Kategoryzacja takich aktywacji mózgu nie jest łatwa bo symboliczne (fonologiczne) reprezentacje pojęć realizowane przez obszary skroniowe reagują na specyficzne aktywacje korowe.
Ostatnie prace (Majjd i inn 2018) pokazują duże zróżnicowanie dokładności werbalizacji różnych zmysłów: w populacji anglojęzycznej ranking dokładności kodowania, od najsłabszej do najlepszej, wygląda tak:
zapach < dotyk < smak < słuch < kształt < kolor.
Jednak w wielu kulturach ta hierarchia wygląda całkiem inaczej, kolejność może być prawie dowolna, zapach może być najlepiej kodowany a nazywanie kształtów bardzo słabo.
W niektórych kulturach afrykańskich zwraca się wielką uwagę na sposób poruszania się i utrzymanie równowagi, istotne w czasie tańców. W języku Anlo-Ewe używanym przez plemiona na południowym wschodzie Ghany jest ponad pięćdziesiąt różnych wyrazów na określenie sposobu poruszania się. Pojęcie "seselelame" oznacza czuj-czuj-ciało-wewnątrz określa odczucia kinestetyczne. Taki sposób zmysłowego odczuwania świata pokazuje, że zachodni dualizm ciało-umysł jest kulturowo uwarunkowany i niezrozumiały w niektórych kulturach (Guerts 2003). Obserwacja ruchu człowieka pozwala ocenić aspekty a nawet walory moralne danej osoby.
|
O przestrzeni peripersonalnej i sztuce paolitycznej - ciekawy artykuł Mirosława Kocura.
B16.6 Synestezje i Ideastezje
Każdy odczuwa świat w nieco inny sposób.
Silne kojarzenie bodźców zmysłowych o różnych modalnościach to synestezje (Gr. syn = razem + aisthesis = postrzegać); nazwę wymyślił
Francis Galton,
Smak lub poza nazwy, widok lub smak dźwięku, czucie widoku, kolory liter ... wszystkie kombinacje.
Wladimir Nabokov - doskonały przypadek kolorowego słuchu.
Długie aaa - odcień starego drewna, g to wulkanizowana guma, k jest jagodowe ...

Zjawisko rzadkie, dla 2 modalności 1:25 000.
Jest przynajmniej 39 rodzajów synestezji!
Najczęściej: wzrok + słuch w formie leksykalno-kolorowej: litery i liczby widziane i słyszane są w kolorach, ok. 2/3 wszystkich przypadków.
Wrażenie koloru może być wywołane przez widok grafemów, cyfr zegara, dźwięków muzyki, ogólnych dźwięków, fonemów, nut, zapachów, smaku, bólu, osobowość rozmówcy, dotyk, temperaturę, podniecenie, emocje.
Bardzo rzadko w synestezjach występuje dotyk i smak, najrzadziej węch; obszar węchowe są oddalone od innych i mają silniejsze projekcje jednokierunkowe w stronę kory analizującej wrażenia o innych modalnościach.
U kobiet zdarza się kilka razy częściej niż u mężczyzn (3-8 razy w różnych badaniach).
Przejściowe synestezje: wywołane anty-serotoninergiczny środkami halucynogennymi, deprywacją sensoryczną, stymulacją elektryczną lub padaczką skroniową, urazami głowy.
Neuronalne mechanizmy synestezji w niektórych przypadkach mogą wynikać z tego, że pola korowe leżą blisko siebie:
- Obszary związane z przetwarzaniem informacji o kolorze (V4) i VWFA (lewa tylna część zakrętu dolnoskroniowego) przylegają do siebie, więc wzajemne oddziaływanie jest stosunkowo silne.
- Połączenie mowy-smaku może wynikać z wzajemnego oddziaływania kory wyspy, analizującej bodźce smakowe, i przylegającej do tego obszaru kory słuchowej.
- Kombinacja smaku i dotyku może być związana z smakowo-dotykowymi projekcjami.

Czym jest wrażenie? Fala świetlna, wibracja mechaniczna czy pobudzenie chemiczne => podobne impulsy elektryczne.
Wrażenia wzrokowe: generyczne, konkretne kształty spiral, siatek, błyszczących ruchomych kropek, stosunkowo proste pobudzenia układu wzrokowego na poziomie V1.
Kolory dotyczą muzyki i mowy, ale wrażenie może być też zależne od ortografii.
Synestezje nie zawsze są przyjemne: usłyszenie "pięć" wywołuje wrażenie cyfry 5 na szarym tle i gwałtowne skurcze prawej części twarzy.
Obrazowanie mózgu pokazuje silne pobudzenia obszarów kory sprzężonych zmysłów.
Wzmożona aktywność struktur limbicznych, obniżona kory - jak u niemowląt.
Hipotezy:
- Synestezje jako silne sprzężenie kory sensorycznej różnych modalności.
- Synestezje jako postrzeganie podkorowe przed określeniem, z którego wrażeń na zmysły.
Obszary leżące w pobliżu (np. reakcji na kolory i liczby) mogą ulec połączeniu.
Tendencje do utrzymania się takich połączeń mają uwarunkowanie genetyczne, synestezje u bliźniąt jednojajowych są silniej skorelowane.
Elizabeth Sulser z Zurichu ma wyjątkowo silne połączenia pomiędzy korą smakową, słuchową i wzrokową.
Podział informacji zmysłowej prawdopodobnie umożliwia większą precyzję działania.
Teoria rozwojowa: niemowlęta do
4 miesiąca doświadczają synestezji.
Mowa pobudza wówczas potencjały wywołane nie tylko w płatach skroniowych ale i potylicznych.
Rzadkie przypadki: pacjent JP rysuje złożone figury geometryczne, które widzi patrząc na równania matematyczne (B. Brogaard, Neurocase 2013).
Do pewnego stopnia każdy ma synestezje, np. polecenia werbalne lub dźwięki mogą w nas wywołać wyobrażenia wzrokowe.

Wielu ludzi w podobny sposób kojarzy ze sobą zapachy, kolory, wrażenia dotykowe, a nawet kolory i kształty, mamy więc do czynienia z podprogową synestezją.
Przykład: który obrazek nazwiemy
Buba a który Kiki?
Co więcej, synestezje można
wywołać sztucznie.
Czytając teksty w których są kolorowe litery lub znaki wytwarza się skojarzenia pomiędzy kolorem i znakami, wiec przypominanie sobie słów zaczyna wiązać się z kolorami liter/słów.
Przykłady takich kolorowych
tekstów są tu.
To samo można zapewne zrobić kolorując nuty. Jak silne są to wrażenia i czy można je porównać z wrodzoną synestezją? To się dopiero okaże w wyniku dalszych badań. Niestety wymaga to dłuższego treningu.
Syn-aesthesis oznacza "współ-postrzeganie", synestezje są wrażeniami zmysłowymi wywołanymi przez inne wrażenia zmysłowe.
Wrażenia zmysłowe mogą tez wywołać procesy poznawcze, reprezentacje semantyczne pojęć, które mogą pobudzać korę zmysłową. Takie zjawisko nazywa się
ideastezją . Np. kolory czy dzwięki cyfr lub innych znaków mogą nie zależeć od kształtu i formy wizualnej tylko od sensu znaku.
Synestezje nabyte w wyniku mechanicznego uszkodzenia mózgu
Derek Amato, muzyczny geniusz po uszkodzeniu mózgu w wyniku uderzenia.
Alonzo Clemons, rzeżbiarz
Nadludzkie zdolności (superhuman showdown).
Dobra strona o energii dla mózgu oraz indeks glikemiczny, wskazujący na poziom glukozy we krwi w parę 3 godzin po spożyciu pokarmu.
Ssaki są stałocieplne. Termoregulacja wymaga termoreceptorów i mechanizmu regulacji wydzielania ciepła.
Liczne inne funkcje homeostatyczne odbywają się całkowicie poza świadomością.
|
Zadanie:
|
Pytania, na które powinniście znać odpowiedzi po przeczytaniu notatek do tego wykładu: wzrok.
- Co to jest optyczna ataksja i z jakimi uszkodzeniami mózgu się wiąże?
- Co to jest i jak się przejawia asymbolia bólu?
- W jaki sposób czuciowe mapy topograficzne mogą się zmieniać?
- Na czym polega dysmorfia ciała i co może być jej przyczyną?
- Hipoteza Milner-Goodale.
- W jaki sposób układ wzrokowy, czuciowy i ruchowym współpracuje ze sobą?
- Opisać drogi słuchowe.
- Pierwotna kora słuchowa i jej połączenia z innymi obszarami.
- Dlaczego kora oczołodołowa jest bardzo ważna dla percepcji?
- Jaka jest rola wtórnej kory słuchowej?
- Jaka jest rola wyspy i równiny skroniowej.
- Co to jest tinnitus?
- Jakie znasz afazje?
- Opisać afazję nazewniczą (lub "opisać afazję xxx").
- Co to jest efekt Mozarta i czy naprawdę występuje?
- Jakie wyróżniamy działy biomuzykologii?
- Dlaczego mamy muzykę?
- Jaką muzykę preferują zwierzęta?
- Co wiesz o drogach wrażeń smakowych?
- Gdzie mieści się kora smakowa?
- Co to jest zespół smakosza?
- Na czym polega szczególne przetwarzanie informacji węchowej w mózgu?
- Dlaczego nie mamy receptorów węchowych po obu stronach głowy, tak jak uszu?
- Wymienić 3 drogi pobudzeń informacji węchowej.
- Gdzie jest pierwotna a gdzie wtórna kora węchowa? Jaka jest jej rola?
- Co to jest anosmia, hyposmia, dysosmia, kakosmia, parosmia, heterosmia, hyperosmia?
- Co to jest ageuzja, hypogeuzja, dysgeuzja, hypergeuzja?
- Gdzie analizowane są bodźce węchowe?
- Co to jest nerw zerowy? Jaka jest jego hipotetyczna rola u ludzi?
- Czy aromaterapia leczy?
- Gdzie mieści się zmysł równowagi?
- Jakie obszary mózgu muszą współdziałać dla utrzymania równowagi?
- Jak wywołać wrażenia autoskopowe? I co to takiego?
- Co to jest allotopagnozja, autotopagnozja, heterotopagnozja.
- Jak często występują synestezje?
- Dlaczego w niektórych mózgach powstają wrażenia synestetyczne?
- Jak działa zmysł pragnienia?
- Jak działa zmysł głodu?
- Zaburzenia łaknienia.
- Czy ludzie mogą orientować się podobnie do nietoperzy? Jak?